【2.5.1】Emini surface accessibility scale
Description: The calculation was based on surface accessibility scale on a product instead of an addition within the window. The accessibility profile was obtained using the formulae Sn = (n+4+i ) (0.37)^-6 where Sn is the surface probability, dn is the fractional surface probability value, and i vary from 1 to 6. A hexapeptide sequence with Sn greater than 1.0 indicates an increased probability for being found on the surface.
A C D E F G H I K L M N P Q R S T V W Y
0.49 0.26 0.81 0.84 0.42 0.48 0.66 0.34 0.97 0.4 0.48 0.78 0.75 0.84 0.95 0.65 0.7 0.36 0.51 0.76
甲型肝炎病毒(HAV)最近在形态学,流行病学和生物物理特征的基础上被分类为肠道病毒72型,是Picornaviridae家族肠道病毒属的成员。 毒粒是浸染的二十面体,其表面由三种结构蛋白即VP1,VP2和VP3(3,19)组成。 VP1是主要暴露的蛋白(10)。中和单克隆抗体的研究表明,该结构蛋白至少含有部分病毒体中和作用。
Emini等人(5,6)在脊髓灰质炎病毒VP1蛋白上鉴定了几个脊髓灰质炎病毒1型特异性中和抗原性位点。 包含这些位点氨基酸序列的合成肽能够直接诱导病毒中和抗体反应,或者在接种亚免疫原剂量的完整病毒体后引发这种免疫反应
最近克隆了基因组HAV RNA的cDNA,并获得了非结构性(20)和结构性(14)基因区域的部分序列,这些信息的可用性促使人们检查了HAV与脊髓灰质炎病毒之间的结构相关性。 希望对后者的抗原结构的详细了解将对HAV的抗原结构提供深刻见解。
一、VP1氨基酸序列比较。
通过Dayhoff(4)的ALIGN程序比较了HAV和1型脊髓灰质炎病毒(Mahoney株)(12,18)的VP1氨基酸序列的一级序列同源性(数据未显示)。 同源性很小,比对得分(4)为1.93,表明与随机排列的序列进行比对会产生更好的得分的概率为2.5%。 通常认为这种同源性不足以仅基于一级序列在蛋白质之间建立进化关系。 具有不同断裂罚分的比对得出相似的比对分数。 尽管如此,仍对HAV和脊髓灰质炎病毒蛋白进行了临时比对。
通过Chou和Fasman(2)和Garnier等人(7)的方法比较了两种VP1蛋白的预测二级结构,结果表明没有预测的结构相似性(数据未显示)。 例如,通过Garnier等(7)的方法,脊髓灰质炎病毒VP1(1至302)和HAV VP1(5至306)的预测α螺旋位势排列不佳,比对相关系数(r)为-0.22 (R2 = 0.049)。前110个残基中的螺旋电位排列实际上显着为负,r = 0.51(r2 = 0.26)表明可能存在反相关。 Garnier等人(7)的螺旋,延伸结构(片),匝或线圈电势或Chouand Fasman(2)描述的可比较电势,其他可能的比对也很差。 脊髓灰质炎病毒VP1的表达与HAV无明显相似性,无法根据预测的二级结构进行比对
因此,为了增加对由Dayhoff程序进行的临时比对的信心,开发了一种方法,用于根据表面概率指标,通过预测的表面特征比较两个序列。 此方法假定没有明显的内部缺失或插入。 对于给定的氨基酸序列,序列号n的一个点是n-2至n + 3位氨基酸表面表面概率的归一化乘积,使用Janin等人(11)的经验性氨基酸可访问表面概率 ,根据28种蛋白质的结构数据确定蛋白质表面氨基酸的分数概率(0.26至0.9)。表面残基定义为水可接触表面的> 20A^2(2.0nm^2)的残基。 利用这些分数的氨基酸表面概率,可以定义序列位置n的表面概率(S)如下:

当通过这种表面概率方法比较两个序列时,发现了惊人的相似性(图1)。这些相似性,结合Dayhoff程序分析的结果,可以对两个VP1序列进行比对,并可以鉴定三个HAV位点, 它们分别对应于先前确定的三个(6)脊髓灰质炎病毒中和抗原位点(图2),一旦对齐,在脊髓灰质炎病毒和HAV位点之间的氨基酸序列和含量相似性就很明显。


二、HAV特异性合成肽诱导抗HAV反应
用Beckman 990B型肽合成仪通过固相方法(1)制备合成肽,该肽包含三个靶向HAV位点之一的氨基酸序列(图3)。 通过免疫吸附分析发现完整的HAV或十二烷基硫酸钠破坏的HAV与游离肽结合(数据未显示)。因此,该肽可能表达与完整病毒体相同的构象或构象集,尽管 交叉反应的性质尚不完全清楚。

然后将该肽独立地通过任一末端共价连接至牛血清白蛋白载体。每次接种时,分别向新西兰白兔和Hartley豚鼠接种0.7 mg相应的结合物。 每只动物在2个月内接受3次弗氏佐剂(Freund adjuvant)注射。 产生的血清均含有抗肽免疫球蛋白G抗体。此外,每种血清还含有免疫球蛋白G抗体,该抗体能够(i)直接结合纯化的甲型肝炎病毒粒子并(i)中和病毒在细胞培养物中的感染性(表1)。 通过暴露于游离肽可以从血清中去除抗HAV抗体(图4)。
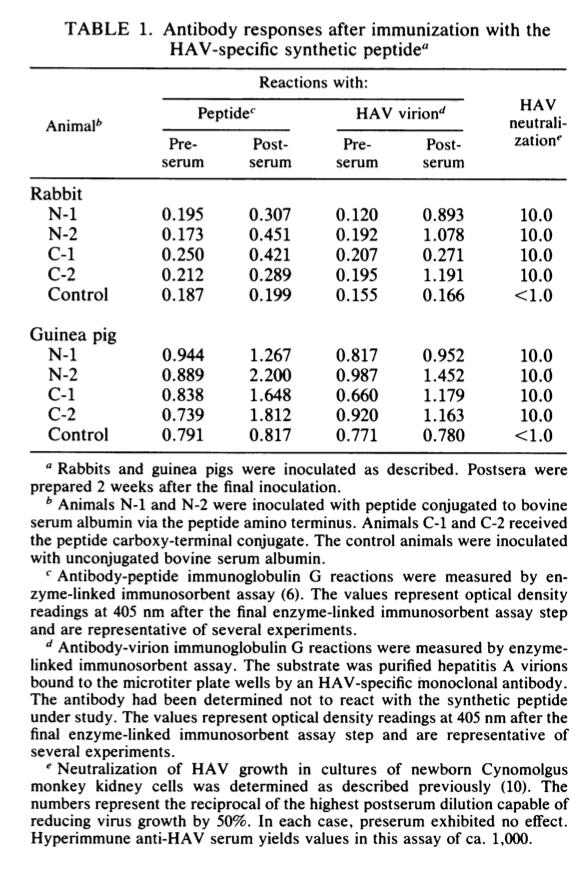
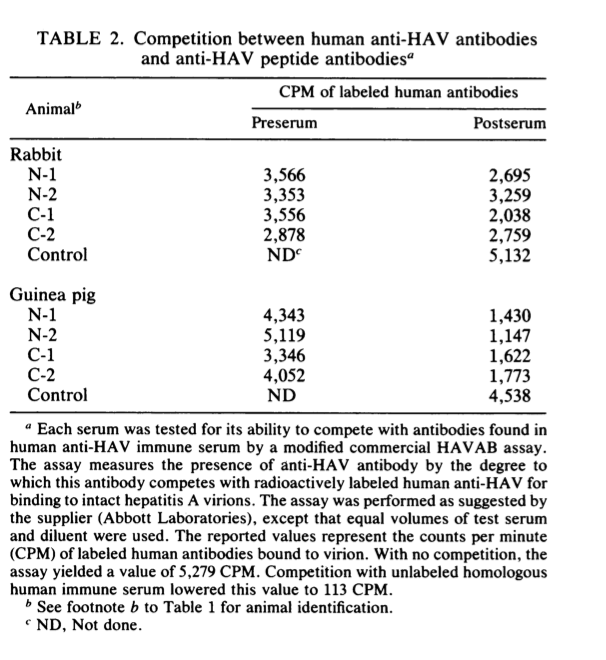
确定该肽诱导的抗病毒颗粒抗体是否也能够与人HAV免疫血清中发现的抗体竞争具有重要意义。为此,每种血清均在改良的商业HAVAB分析中进行了测试,该方法通过与 标准的人类免疫血清。 结果(表2)表明,所有的豚鼠和50%的兔血清都含有这种竞争性抗体。 因此,一些交叉反应的,肽特异性的病毒体表位也可以被人的免疫系统识别。但是,值得注意的是,至少一个被兔免疫系统识别的表位未被人识别。
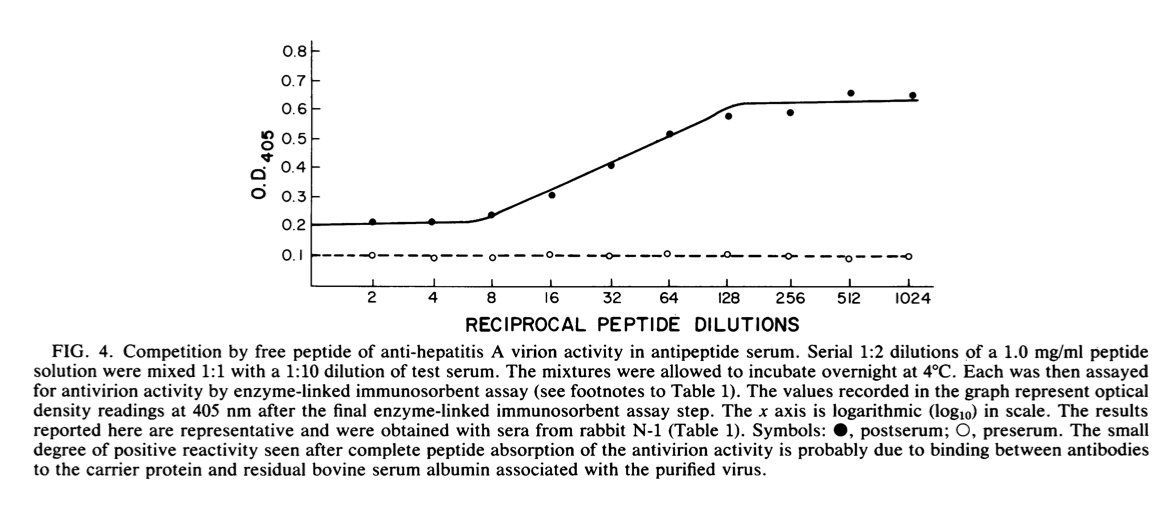
最后,通过Western免疫印迹发现抗肽抗体与纯化的变性HAV VP1反应(数据未显示)。该结果强调了病毒体表面上肽序列代表的决定簇的潜在顺序性质。
令人感兴趣的是,尽管看似最小的一级序列和二级结构保守性,脊髓灰质炎病毒和HAV之间的表面结构同源性可能是显着的。 这种结构相似性可能涉及两种病毒的抗原性位点呈递。 显然,仅通过与脊髓灰质炎病毒特异性VP1上此类抗原性位点的已知位置进行比较即可确定HAV特异性中和性抗原性位点。 该通报中报道的合成肽作为合成HAV疫苗的组成部分具有巨大潜力,尤其是因为已显示抗肽抗体可与人HAV免疫血清中的抗体竞争。 目前正在研究含有其余两个可能的HAV中和抗原位点氨基酸序列的合成肽。
同样,总的来说,应该指出的是,通过表面概率分析对HAV和脊髓灰质炎病毒VP1序列进行比对,证实了当前HAV VP1在HAV基因组RNA序列上的定位。该定位是通过将RNA序列与VP1进行比较而建立的 氨基酸序列数据(14)。
参考资料
- Emini EA, Hughes JV, Perlow DS, Boger J. Induction of hepatitis A virus-neutralizing antibody by a virus-specific synthetic peptide. J Virol. 1985 Sep;55(3):836-9.
- http://tools.iedb.org/bcell/help/
