【3.2.4】几种不常见的DNA二级结构
1. DNA弯曲(Bending)
DNA 的双螺旋结构是相对稳定的,有时会出现弯曲。弯曲的形成一方面与一些特殊的碱基序列有关,另一方面与某些DNA结合蛋白的作用有关。DNA的弯曲可以通过电泳的方法检测到。例如,一种原生动物的动体DNA(kinetoplast DNA,kDNA)的实际长度为414bp,但其电泳的行为显示它有828bp。正是弯曲降低了它的泳动速率!碱基序列分析显示,含有成串A重复序列(4-5个A)且每串A前后分别是C和T的片段之间容易形成弯曲,如 CCC (5A) TCTC (6A) TAGGC (6A) TGCC (5A) TCCCAAC。这是因为成串A构成的片段因碱基堆积力较强形成更稳定的双螺旋,这里的双螺旋与标准的B型双螺旋有所变化,如1圈只有10bp,具有更窄的小沟,但在它们之间的序列却易出现弯曲。
在基因转录或激活的时候,可观察到DNA上游序列发生弯曲。那些参与调节基因表达的蛋白质虽然结合在距离基因较远的地方,但通过弯曲DNA,而拉近与其他序列之间的距离。弯曲的DNA片段可能位于基因的上游、下游和启动子附近。此外,DNA弯曲还发现在DNA复制、位点特异性重组和DNA修复过程中。DNA弯曲也有利于压缩比较大的基因组DNA。
已发现类似CTGnCAGn 的序列比其他序列更容易组装成核小体的结构,这是因为这样的序列构成的双螺旋具有较好的柔性,容易发生弯曲。这说明DNA弯曲可影响到染色质结构的组装。

2. 十字形DNA(Cruciforms)
这种结构的形成需要特殊的碱基序列,即反向重复序列(inverted sequence)。当2段长于10bp的反向重复序列被一小段非重复序列隔开以后,它们可以通过链内的互补碱基对同时在两条链上形成2个对称的发夹结构,结合在一起像十字形。这种结构可能作为某些转录因子结合的位点,从而参与特定基因表达的调控。

3. 三螺旋DNA (Triple helix)
双螺旋的poly d(A)-poly d(T)在一定条件下可以容纳第三条poly d(T)链。第三条链存在于双螺旋的大沟,通过与嘌呤形成碱基对而与双螺旋形成稳定的结构(图2-25)。采用核磁共振技术研究发现,当DNA溶液中poly嘌呤和poly嘧啶的化学计量比为1:2时形成的poly核苷酸链,可以形成三链DNA。在低pH条件下,如果DNA序列包含长的poly(嘌呤)-poly(嘧啶),在DNA分子内部也可形成这种三链结构,这种构象的DNA被称为H-DNA。在细胞内富含嘧啶的DNA链与其对应的DNA链部分解离,向后折叠,结合在Watson–Crick双螺旋的大沟内,与富含嘌呤的DNA链形成氢键。这种结构的形成对基因表达起调控作用。
这种结构的形成涉及到3段碱基序列,每一段序列要么全是嘌呤,要么全是嘧啶,而且具有互补的关系。可以是两段相同的全嘌呤序列和一段互补的全嘧啶序列,也可以是两段相同的全嘧啶序列和一段互补的全嘌呤序列。三螺旋结构的形成可以影响到DNA的复制、转录,还可能阻止特定的蛋白质和DNA的结合。
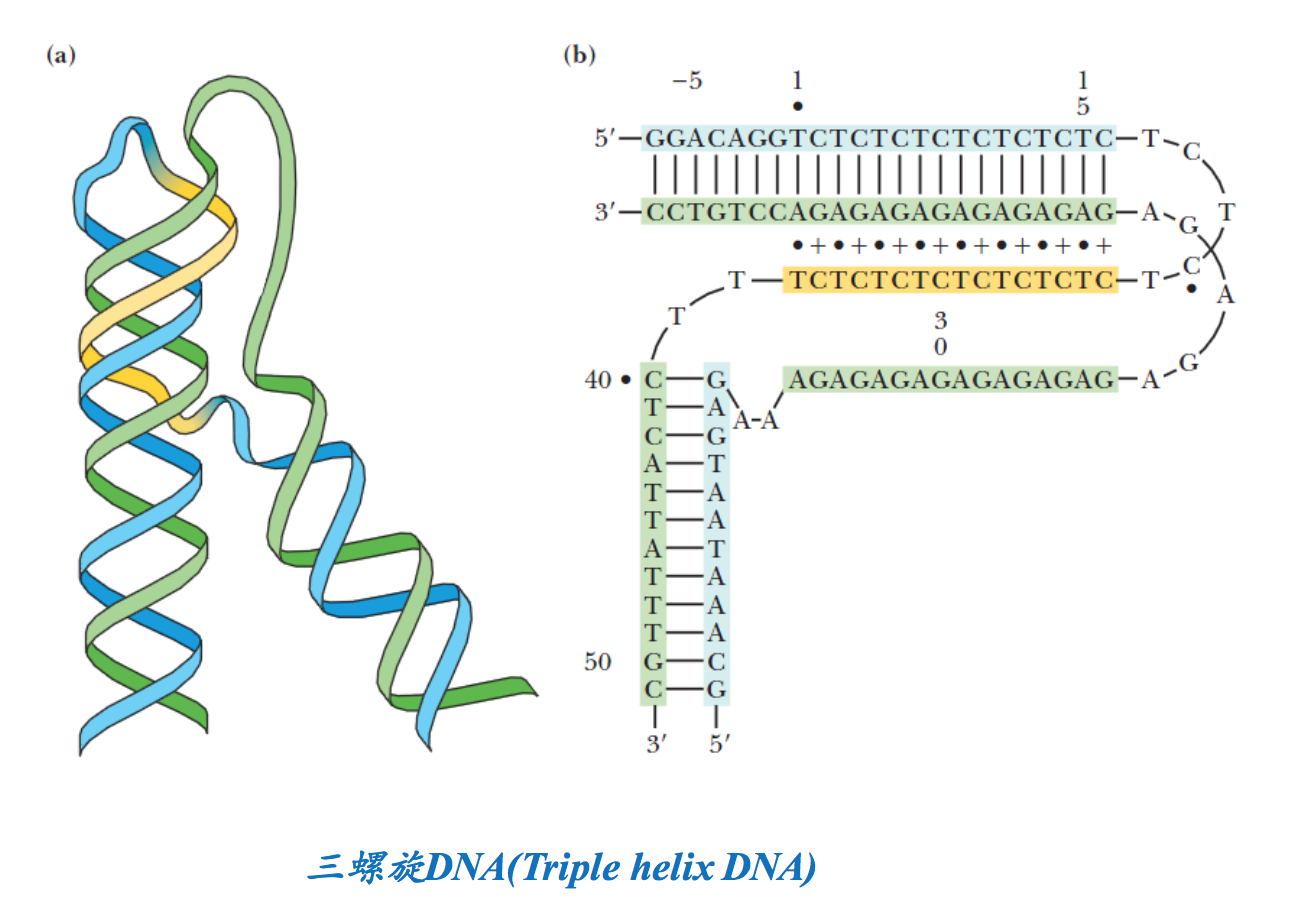
三螺旋结构可以在两个DNA分子之间形成,也可能在同一个DNA分子内形成。前者容易在两个DNA分子上全是嘌呤和全是嘧啶的互补区段之间形成,但第三条链在配对结合的时候有两种不同的方向,而具体的方向取决于链的性质。第三条链通过Hoogsteen氢键配对(表2-3)的时候,需要C的质子化(低pH)。这样的三螺旋结构可能在DNA同源重组的时候形成:一个DNA分子上的同源片段出现裂口,然后发生解链,其中的一条链与另一个DNA分子上的同源双链配对结合。
分子内的三螺旋结构形成,除了需要互补的全嘌呤和全嘧啶序列以外,还需要序列呈镜像重复。第三条链上的C需要质子化才能与G配对。已发现,超螺旋的形成有利于这类三螺旋结构的形成。
4. 四链DNA (G-quadruplex)
很长一段时间内,没有人敢想过DNA能形成四链结构。但在三螺旋结构被发现以后,有人根据Watson-Crick碱基对和Hoogseen碱基对,首先从理论上预测出四链结构是可能存在的。很快在体外发现,由CGG重复序列组成的单链DNA在K+、Na+或Li+存在下很容易形成四链结构,这种结构可使用凝胶电泳的方法检测出来,因为它泳动的速度比其他形式的DNA要快! 化学修饰实验清楚地显示G参与以Hoogsteen氢键形成四链。在两个两侧含有2段GGGG(G四联体,G quartets)重复序列的GCGC序列组成的DNA片段之间,可以形成四链结构.
真核生物染色体DNA的端粒是细胞内DNA最可能形成四链结构的地方!端粒DNA是由短的GnTn重复序列组成。重复的次数成百上千。如四膜虫大核端粒DNA重复序列GGGGTT,拟南芥为GGGATTT,人类为GGGATT。
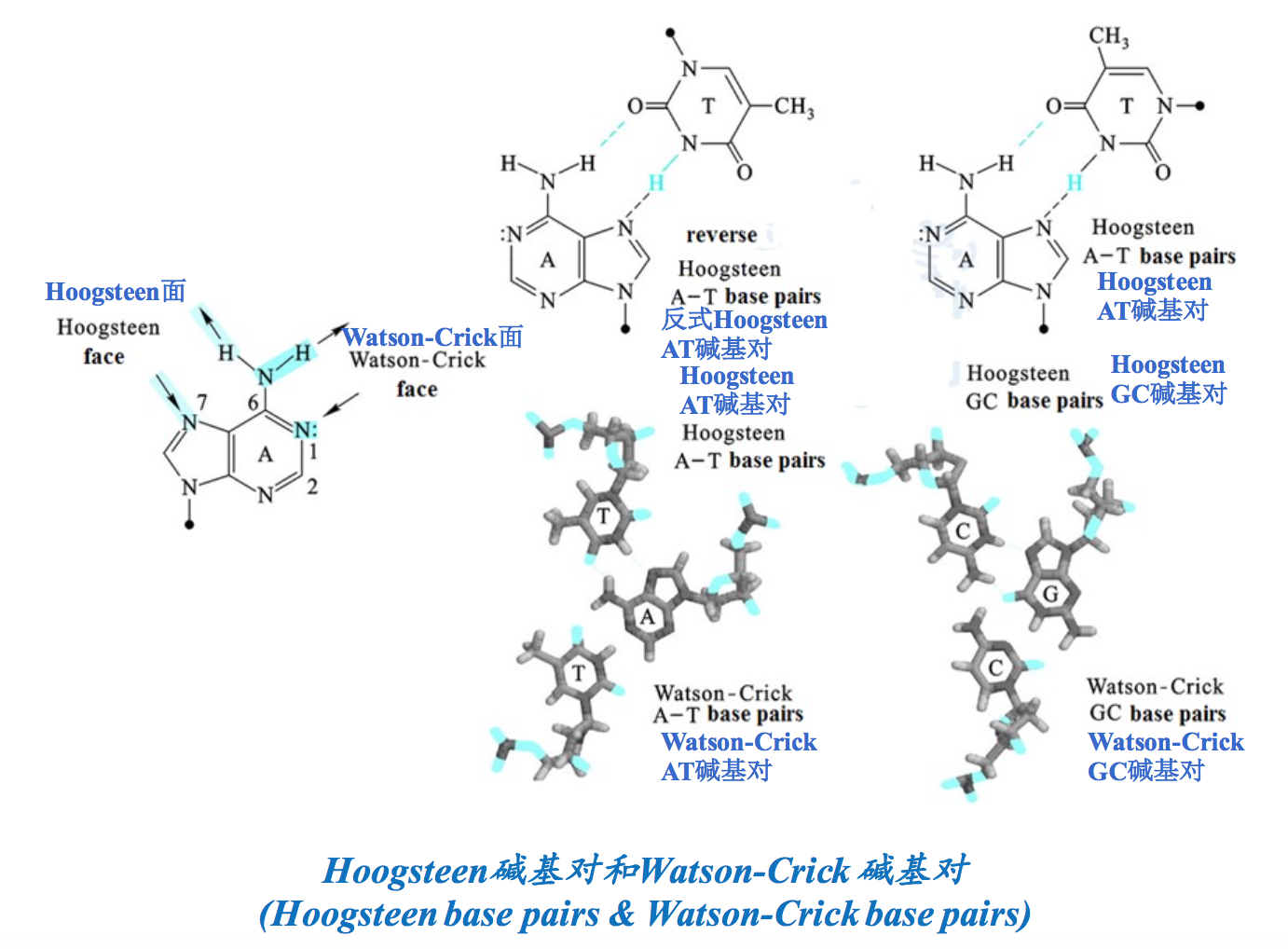
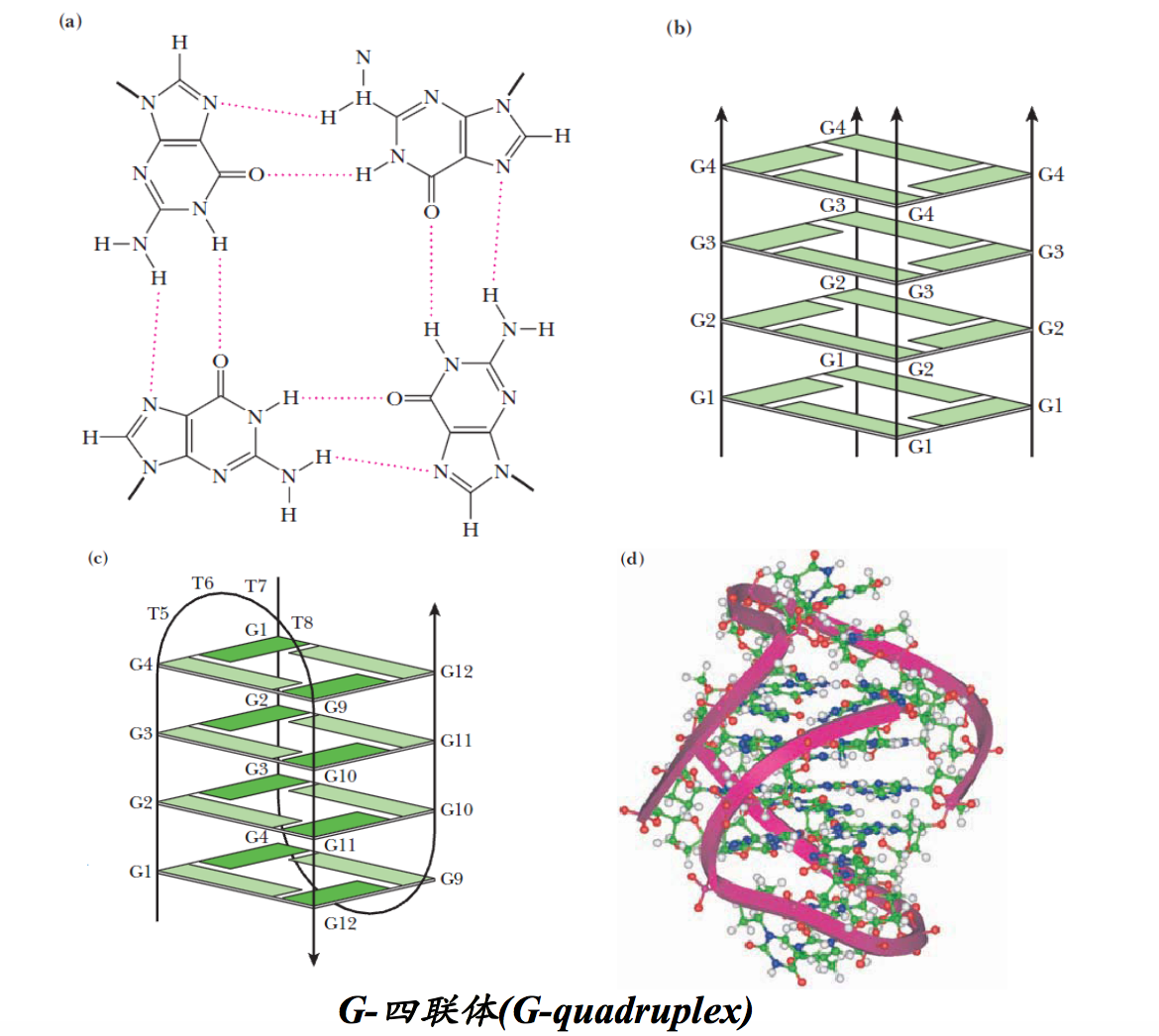
多数真核细胞的端粒DNA的3′-端具有由富含G的短重复序列组成的悬垂(overhang),长度通常为12~16个碱基。人端粒DNA的3′-端的悬垂较长,有125~275个碱基,其重复序列为GGGATT。人工合成的人悬垂序列在体外特定的条件下可以通过G-四联体结构形成四链结构。在G四联体结构之中,每一个G通过Watson-Crick面与相邻G的Hoogsteen面形成氢键。四个G的O6位于四联体的中心,每两个四联体片层可以结合一个金属离子。
四联体可能是平行的,也可能是反平行的。如果是反平行的,相邻的G就必须采取不同的取向。
有人根据体外的一些实验数据认为,人细胞端粒DNA的悬垂能自我环绕,折叠成为螺旋桨状的G-四联体(G-quadruplex),这种结构可与特定的端粒DNA结合蛋白结合,可提供额外的保护,有助于它的完整性和稳定性。此外,体内的四联体可能参与调节c-Myc癌基因的转录:有证据表明,一种椅状的四联体(chair-G-quadruplex)结构能够阻止c-Myc的转录激活。如果四联体被破坏,c-Myc的转录就会显著增加。
在染色体端粒富含G的区域,DNA可形成四链结构,在四个G之间形成Hoogsteen氢键。Poly(G)-DNA链可采用四链平行排列的方式形成四链DNA构象,具有A/T间隔富含G的DNA序列两条链反向折叠形成分子内部的四链DNA构象。
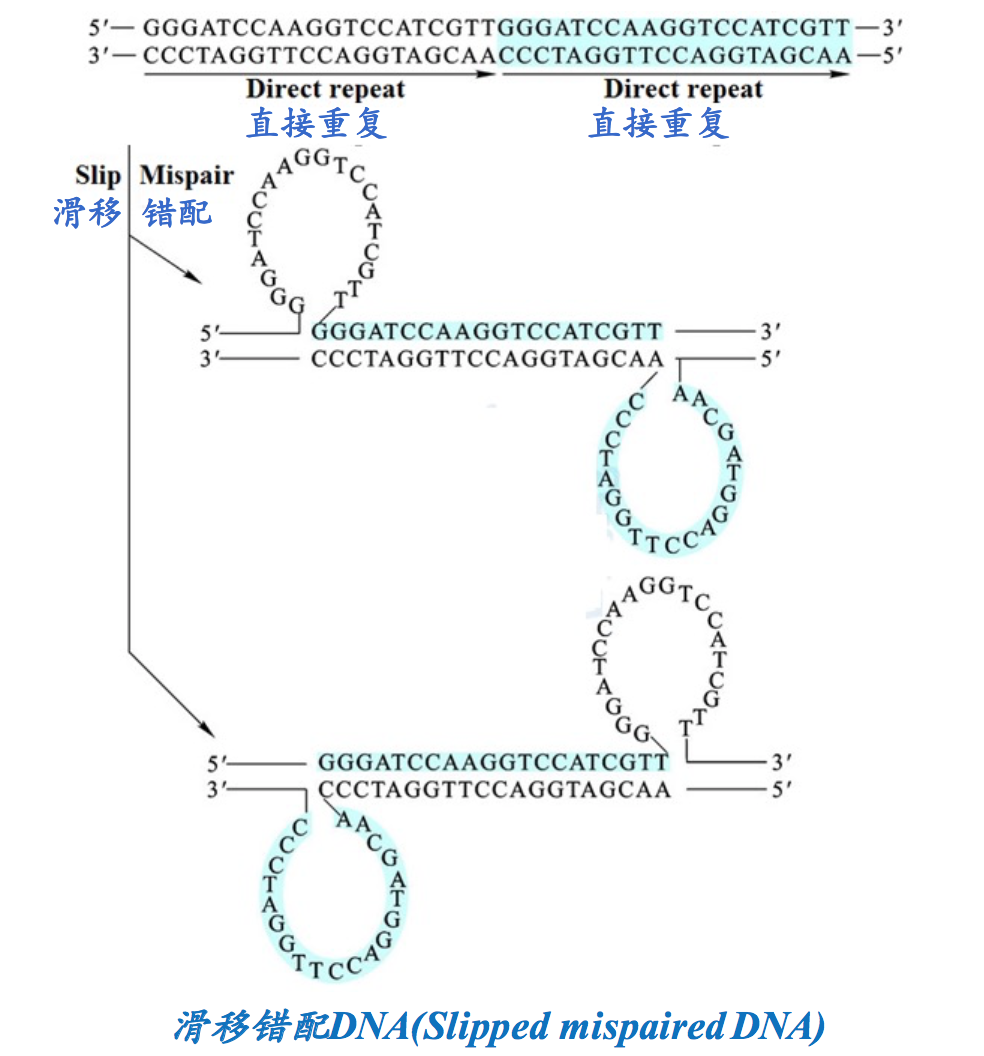
5. 错配滑移DNA (Slipped mispaired DNA, SMP-DNA)
滑移错配DNA。含有直接重复序列的DNA可以形成一种叫“滑移错配”的二级结构。形成这种结构的原因是该区段DNA先发生解链,在重新缔合的时候,一段重复单元内的核苷酸序列因滑移与另一段重复单元内的互补序列发生错配,从而形成两个环。因滑移的方式不同,可形成两种SMP-DNA。若体内的DNA形成上述结构,会导致某些基因发生移框突变。
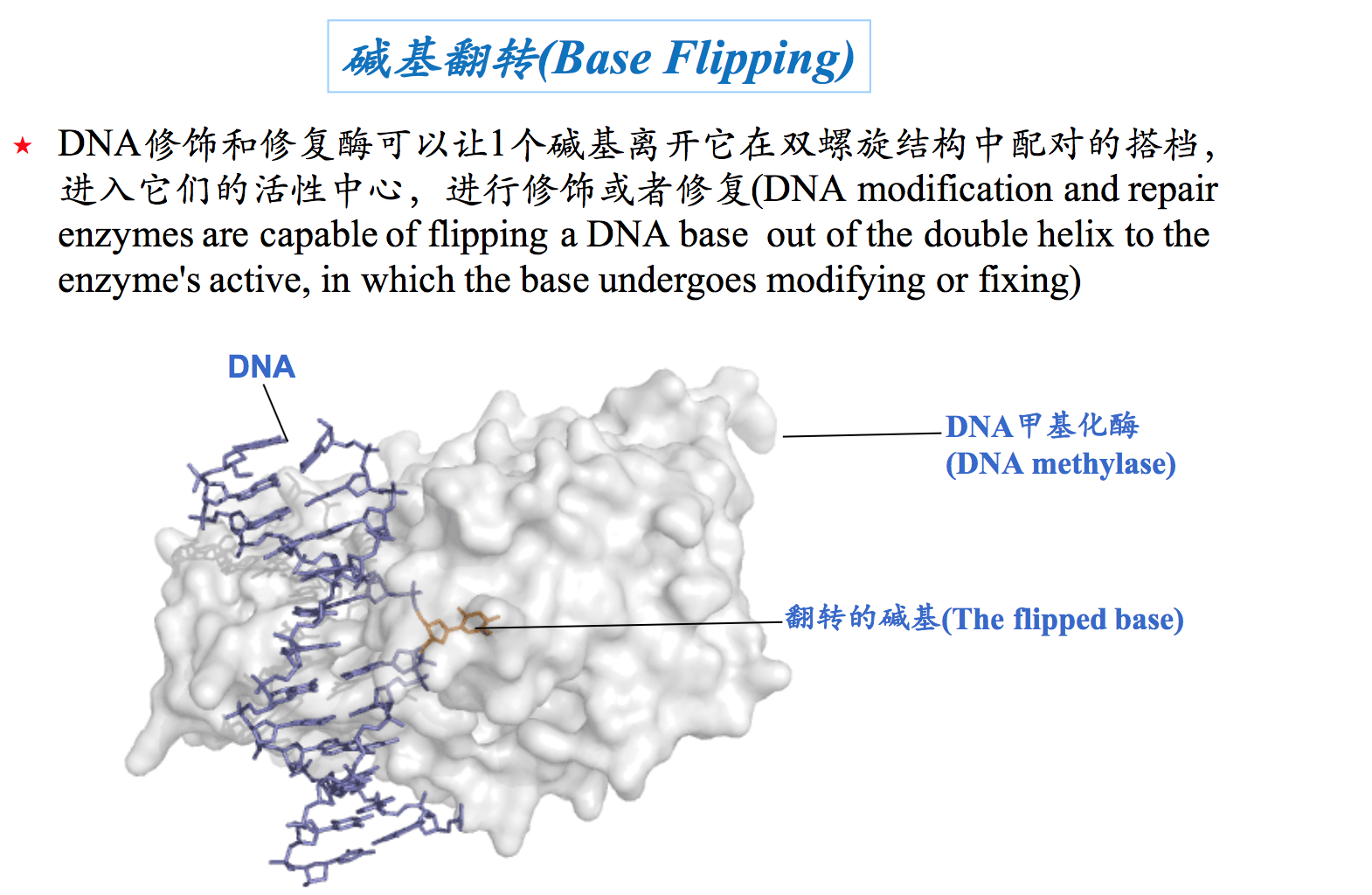
6. 碱基翻转 (Base flipping)
有时候,DNA双螺旋上的某个碱基离开它的“配偶”,突出在双螺旋之外,这种现象称为碱基翻转。一个碱基发生翻转的时候,会造成相邻碱基对的扭曲。碱基翻转对于细胞的某些功能是很重要的。例如,参与同源重组的酶需要通过碱基翻转寻找同源的序列,催化碱基修饰的酶需要碱基通过翻转落入它的活性中心被化学修饰,参与碱基切除修复的DNA糖苷酶需要受损伤的碱基通过翻转进入它的活性中心被切除。最后一种情况的实例就是,DNA分子上出现的U是通过这种方式被尿嘧啶-DNA糖苷酶切除掉的。
参考资料
- 南京大学 杨荣武老师 《结构生物学》课件
