【3.6.1.1】蛋白质折叠和加工
翻译完成了细胞内遗传信息的流动。DNA中的核苷酸序列现在已经转化为多肽链中的氨基酸序列。然而,多肽的合成并不等同于功能蛋白的生产。为了有用,多肽必须折叠成不同的三维构象,在许多情况下,多个多肽链必须组装成功能复合物。此外,许多蛋白质经过进一步的修饰,包括碳水化合物和脂质的切割和共价连接,这对蛋白质在细胞内的功能和正确定位至关重要。
一、伴侣与蛋白质折叠
蛋白质的三维构象是由其组成氨基酸的侧链之间的相互作用产生的,如第2章所述。蛋白质折叠的经典原理是,蛋白质采用正确的三维构象所需的所有信息都由其氨基酸序列提供。这一点最初是由Christian Anfinsen的实验证明的,变性的RNase可以在体外自发地重折叠到其活性构象(见图2.17)。因此,蛋白质折叠似乎是一个自组装过程,不需要额外的细胞因子。然而,最近的研究表明,这并不能充分描述蛋白质在细胞内的折叠。蛋白质在细胞内的正确折叠是由其他蛋白质的活性介导的。
促进其他蛋白质折叠的蛋白质被称为分子伴侣(molecular chaperones.)。“伴侣”一词最初由Ron Laskey和他的同事用来描述一种蛋白质(核浆蛋白),它是从组蛋白和DNA组装核小体所必需的。核浆蛋白与组蛋白结合并介导它们组装成核小体,但核浆蛋白本身并没有结合到最终的核小体结构中。因此,伴侣作为催化剂促进组装,而不是组装复合体的一部分。随后的研究将这一概念扩展到包括介导各种其他组装过程的蛋白质,特别是蛋白质折叠。
需要注意的是,伴侣蛋白不会传递多肽折叠成其正确三维构象所需的额外信息;蛋白质的折叠构象仅由其氨基酸序列决定。相反,伴侣通过协助自组装过程来催化蛋白质折叠。它们似乎通过结合和稳定未折叠或部分折叠的多肽发挥作用,这些多肽是通往最终正确折叠状态的途径中的中间体。在缺乏伴侣的情况下,未折叠或部分折叠的多肽链在细胞内会不稳定,经常折叠错误或聚集成不溶性复合物。伴侣蛋白的结合稳定了这些未折叠的多肽,从而防止了错误的折叠或聚集,并使多肽链折叠成正确的构象。
一个很好的例子是伴侣蛋白与仍在核糖体上翻译的新生多肽链结合,从而在链合成完成之前防止多肽氨基末端部分的错误折叠或聚集(图7.17),这种相互作用对于羧基末端(最后一个要合成的)是正确折叠氨基末端所必需的蛋白质来说尤其重要。在这种情况下,伴侣结合使氨基末端部分稳定在未折叠构象,直到多肽链的其余部分被合成,并且完整的蛋白质可以正确折叠。伴侣蛋白在转运到亚细胞细胞器的过程中也稳定未折叠的多肽链,例如,在蛋白质从胞质溶胶转移到线粒体的过程中(图7.18)。蛋白质以部分未折叠的构象转运通过线粒体膜,这些构象由胞质溶胶中的伴侣蛋白稳定。线粒体内的伴侣促进多肽链通过膜的转移及其随后在细胞器内的折叠。此外,伴侣蛋白参与由多个多肽链组成的蛋白质的组装,参与大分子结构(如核纤溶酶)的组装,以及(如本章稍后所述)参与蛋白质降解的调节。
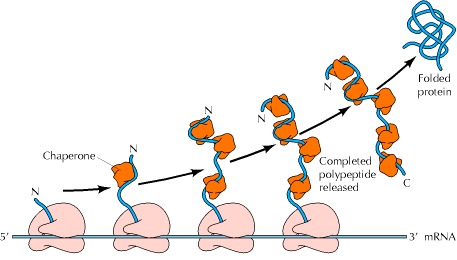
Figure 7.17Action of chaperones during translation
Chaperones bind to the amino (N) terminus of the growing polypeptide chain, stabilizing it in an unfolded configuration until synthesis of the polypeptide is completed. The completed protein is then released from the ribosome and is able to fold into its correct three-dimensional conformation.
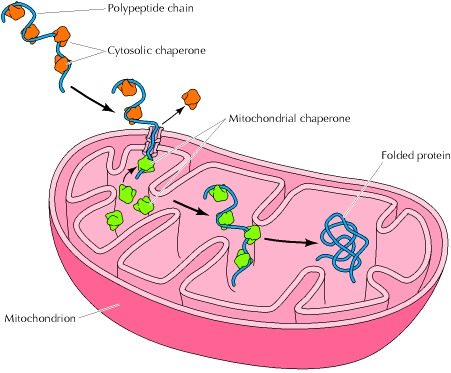
Figure 7.18Action of chaperones during protein transport
A partially unfolded polypeptide is transported from the cytosol to a mitochondrion. Cytosolic chaperones stabilize the unfolded configuration. Mitochondrial chaperones facilitate transport and subsequent folding of the polypeptide chain within the organelle.
许多现在已知具有分子伴侣功能的蛋白质(表7.2)最初被鉴定为热休克蛋白,这是一组在经历高温或其他形式的环境应激的细胞中表达的蛋白质。热休克蛋白(缩写Hsp)在原核细胞和真核细胞中都是高度保守的,被认为可以稳定和促进因暴露于高温而部分变性的蛋白质的重折叠。然而,热休克蛋白家族的许多成员在正常生长条件下表达并具有必要的细胞功能。这些蛋白质充当分子伴侣,在正常条件下以及在受到环境胁迫的细胞中,多肽折叠和运输都需要分子伴侣。
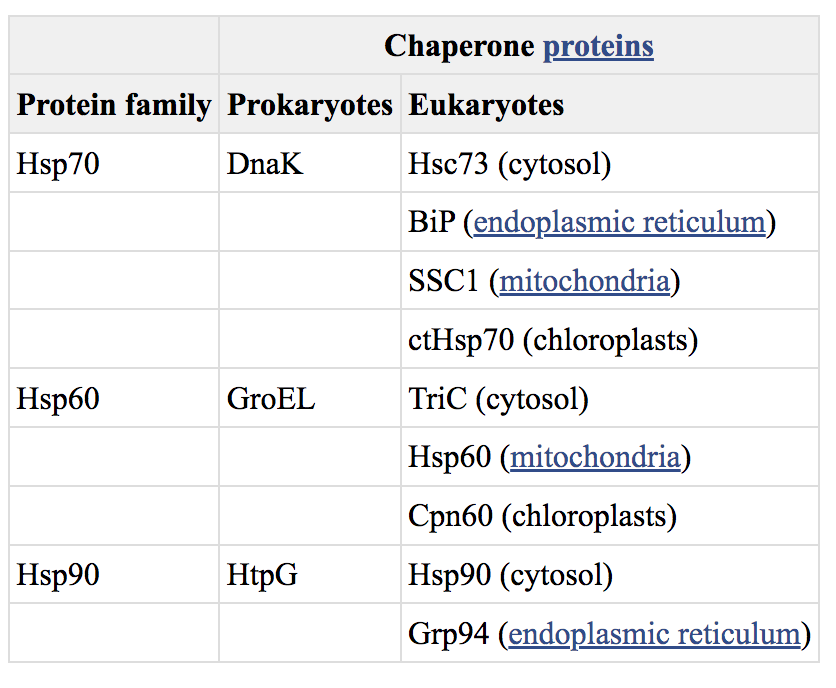
Table 7.2Molecular Chaperones
热休克蛋白的Hsp70和Hsp60家族在原核细胞和真核细胞中的蛋白质折叠的一般途径中似乎特别重要。这两个家族的蛋白质通过与多肽链的未折叠区域结合而发挥作用。Hsp70家族成员在翻译过程中稳定未折叠的多肽链(例如,见图7.17),以及在多肽转运到各种亚细胞区室(如线粒体和内质网)的过程中。这些蛋白质与未折叠多肽的短片段(七或八个氨基酸残基)结合,使多肽链保持未折叠构型并防止聚集。
Hsp60家族成员(也称为伴侣蛋白)促进蛋白质折叠成其天然构象。每个伴侣蛋白由14个亚基组成,每个亚基约60千道尔顿(kd),排列在两个堆叠的环中,形成“双甜甜圈”结构(图7.19)。未折叠的多肽链通过结合在伴侣蛋白圆柱体的中心腔内而与胞质溶胶屏蔽。在这种分离的环境中,蛋白质折叠可以进行,而多肽链的未折叠片段的聚集通过其与伴侣蛋白的结合而被阻止。未折叠多肽与伴侣蛋白的结合是一种可逆反应,与ATP作为能量来源的水解偶联。因此,ATP水解驱动多轮释放并将多肽链的未折叠区域重新结合到伴侣蛋白上,使多肽逐渐折叠成正确的构象。
Figure 7.19
Structure of a chaperonin. GroEL, a member of the Hsp60 family, is a porous cylinder composed of two stacked rings. Each ring consists of seven subunits. (Courtesy of Paul B. Sigler, Yale University.)
在某些情况下,已经发现Hsp70和Hsp60家族的成员以顺序的方式共同作用。例如,Hsp70和Hsp60家族成员在蛋白质转运到线粒体和大肠杆菌中新合成蛋白质的折叠过程中顺序发挥作用(图7.20)。首先,Hsp70 伴侣稳定新生多肽链,直到蛋白质合成完成。然后将未折叠的多肽链转移到Hsp60伴侣蛋白上,蛋白质在其中折叠,产生正确折叠成其功能性三维构象的蛋白质。Hsp70和Hsp60家族的成员存在于真核细胞的胞质溶胶、亚细胞细胞器(如线粒体)以及细菌中(见表7.2),因此Hsp70与Hsp60的顺序作用似乎代表了蛋白质折叠的一般途径。胞质溶胶和内质网中某些蛋白质折叠的另一种途径可能涉及Hsp70和Hsp90家族成员的顺序作用,尽管Hsp90的功能尚不清楚。
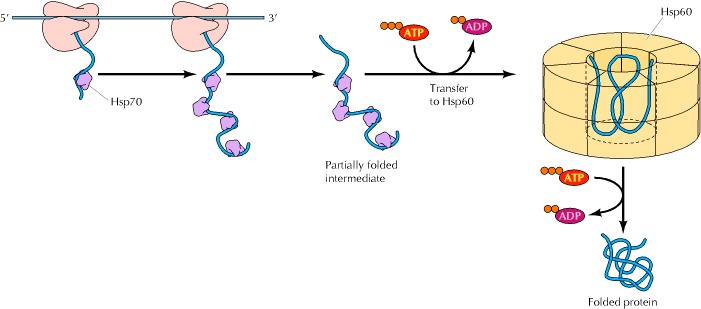
Figure 7.20Sequential actions of Hsp70 and Hsp60 chaperones
Chaperones of the Hsp70 family bind to and stabilize unfolded polypeptide chains during translation. The unfolded polypeptide is then transferred to chaperones of the Hsp60 family, within which protein folding takes place. ATP hydrolysis is required for release of the unfolded polypeptide from Hsp70 as well as for folding within Hsp60.
二、酶与蛋白质折叠
除了通过结合和稳定部分折叠的中间体来促进蛋白质折叠的伴侣蛋白外,细胞还含有至少两种通过破坏和重新形成共价键来催化蛋白质折叠的酶。半胱氨酸残基之间二硫键的形成对稳定许多蛋白质的折叠结构很重要(见图2.16)。Christian Anfinsen于1963年发现的蛋白质二硫异构酶催化这些键的断裂和重新形成(图7.21)。对于含有多个半胱氨酸残基的蛋白质,蛋白质二硫化物异构酶(PDI)通过促进成对二硫化物之间的快速交换发挥着重要作用,从而使蛋白质获得与其稳定折叠构象相容的二硫键模式。二硫键通常仅限于分泌蛋白和一些膜蛋白,因为胞质溶胶含有还原剂,可以将半胱氨酸残基保持在还原的(-SH形式),从而防止二硫键的形成。 在真核细胞中,二硫键在内质网中形成,内质网中维持着氧化环境。与二硫键在稳定分泌蛋白中的作用一致,内质网中PDI的活性与不同类型细胞中的蛋白质分泌水平相关。
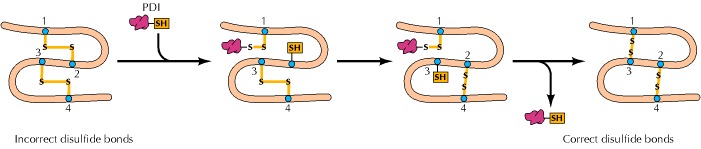
Figure 7.21The action of protein disulfide isomerase
Protein disulfide isomerase (PDI) catalyzes the breakage and rejoining of disulfide bonds, resulting in exchanges between paired disulfides in a polypeptide chain. The enzyme forms a disulfide bond with a cysteine residue of the polypeptide and then exchanges its paired disulfide with another cysteine residue. In this example, PDI catalyzes the conversion of two incorrect disulfide bonds (1-2 and 3-4) to the correct pairing (1-3 and 2-4).
在蛋白质折叠中起作用的第二种酶催化涉及脯氨酸残基的肽键异构化(图7.22)。脯氨酸是一种不寻常的氨基酸,因为脯氨酸残基之前的肽键的顺式和反式构象之间的平衡仅略微有利于反式形式。相比之下,其他氨基酸之间的肽键几乎总是反式的。脯氨酸肽键的顺式和反式构型之间的异构化是由肽基-脯氨酸异构酶催化的,否则这可能代表蛋白质折叠的限速步骤。这种酶广泛分布于原核细胞和真核细胞中,可以催化至少一些蛋白质的重折叠。然而,它在生理上的重要底物和在细胞内的作用尚未确定。
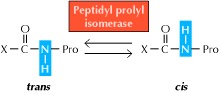
Figure 7.22 The action of peptidyl prolyl isomerase. Peptidyl prolyl isomerase catalyzes the isomerization of peptide bonds that involve proline between the cis and trans conformations.
三、蛋白质切割
多肽链的裂解(蛋白水解)是许多蛋白质成熟的重要步骤。一个简单的例子是从许多多肽的氨基末端去除起始甲硫氨酸,这发生在生长的多肽链的氨基末端从核糖体中出现后不久。另外的化学基团,例如乙酰基或脂肪酸链(稍后讨论),然后经常添加到氨基末端残基。
氨基末端的蛋白质水解修饰也在许多蛋白质跨膜易位中发挥作用,包括细菌和真核生物中的分泌蛋白质,以及旨在结合到真核细胞质膜、溶酶体、线粒体和叶绿体中的蛋白质。这些蛋白质通过氨基末端序列被靶向运输到目的地,当蛋白质穿过膜时,氨基末端序列通过蛋白水解切割被去除。例如,氨基末端信号序列,通常约20个氨基酸长,在翻译仍在进行时,将分泌蛋白靶向细菌质膜或真核细胞内质网(图7.23)。该信号序列主要由疏水性氨基酸组成,从核糖体中出来时插入膜中。随着翻译的进行,多肽链的剩余部分通过膜中的通道。然后,信号序列被特定的膜蛋白酶(信号肽酶)切割,成熟蛋白被释放。在真核细胞中,生长中的多肽链易位到内质网是靶向蛋白质分泌、掺入质膜或掺入溶酶体的第一步。第9章和第10章将详细讨论将蛋白质转运到这些目的地的机制,以及其他靶向序列在将蛋白质导入线粒体和叶绿体中的作用。
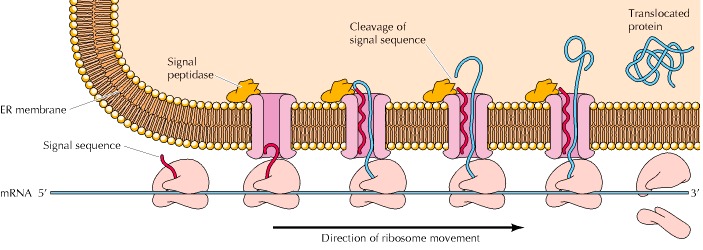
Figure 7.23The role of signal sequences in membrane translocation
Signal se-quences target the translocation of polypeptide chains across the plasma membrane of bacteria or into the endoplasmic reticulum of eukaryotic cells (shown here). The signal sequence, a stretch of hydrophobic amino acids at the amino terminus of the polypeptide chain, inserts into a membrane channel as it emerges from the ribosome. The rest of the polypeptide is then translocated through the channel and the signal sequence is cleaved by the action of signal peptidase, releasing the mature translocated protein.
在蛋白水解加工的其他重要实例中,活性酶或激素通过切割较大的前体而形成。胰岛素,作为一种较长的前体多肽合成,就是一个很好的例子。胰岛素通过两次裂解形成。最初的前体(胰岛素原)包含一个氨基末端信号序列,将多肽链靶向内质网(图7.24)。在转移到内质网的过程中去除信号序列会产生第二个前体,称为胰岛素原。然后,通过蛋白水解去除内部肽,将这种前体转化为胰岛素,胰岛素由通过二硫键连接在一起的两条链组成。其他被类似切割过程激活的蛋白质包括消化酶和参与血液凝固的蛋白质。

Figure 7.24Proteolytic processing of insulin
The mature insulin molecule consists of two polypeptide chains (A and B) joined by disulfide bonds. It is synthesized as a precursor polypeptide (preproinsulin) containing an aminoterminal signal sequence that is cleaved during transfer of the growing polypeptide chain to the endoplasmic reticulum. This cleavage yields a second precursor (proinsulin), which is converted to insulin by further proteolysis, removing the internal connecting polypeptide.
值得注意的是,许多动物病毒的蛋白质来源于较大前体的切割。HIV提供了蛋白水解在病毒复制中作用的一个特别重要的例子。在HIV的复制过程中,病毒编码的蛋白酶切割前体多肽形成病毒结构蛋白。由于其在病毒复制中的核心作用,HIV蛋白酶(除逆转录酶外)是开发治疗艾滋病药物的重要靶点。事实上,这种蛋白酶抑制剂现在是对抗这种疾病最有效的药物之一。
四、糖基化
许多蛋白质,特别是真核细胞中的蛋白质,通过添加碳水化合物进行修饰,这一过程称为糖基化。添加了碳水化合物链的蛋白质(称为糖蛋白)通常被分泌或定位在细胞表面,尽管一些细胞核和胞质蛋白质也被糖基化。糖蛋白的碳水化合物部分在内质网中的蛋白质折叠、将蛋白质靶向递送到适当的细胞内区室以及作为细胞-细胞相互作用中的识别位点中发挥重要作用。
根据碳水化合物侧链的连接位置,糖蛋白分为N-连接或O-连接(图7.25)。在N-连接的糖蛋白中,碳水化合物连接到天冬酰胺侧链中的氮原子上。在O-连接的糖蛋白中,丝氨酸或苏氨酸侧链中的氧原子是碳水化合物附着的位点。直接连接到这些位置的糖通常分别是N-乙酰葡糖胺或N-乙酰半乳糖胺。
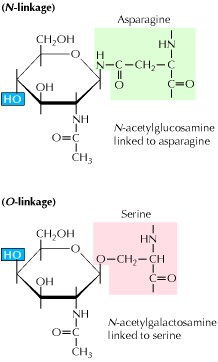
Figure 7.25Linkage of carbohydrate side chains to glycoproteins
The carbohydrate chains of N-linked glycoproteins are attached to asparagine; those of O-linked glycoproteins are attached to either serine (shown) or threonine. The sugars joined to the amino acids are usually either N-acetylglucosamine (N-linked) or N-acetylgalactosamine (O-linked).
真核细胞中的大多数糖蛋白要么用于分泌,要么用于掺入质膜。这些蛋白质通常被转移到内质网(通过信号序列的切割),而它们的翻译仍在进行中。糖基化也在翻译完成之前在内质网中启动。第一步是将由14个糖残基(2个N-乙酰葡糖胺、3个葡萄糖和9个甘露糖)组成的常见低聚糖转移到生长中的多肽链的天冬酰胺残基上(图7.26)。低聚糖在内质网内组装在脂质载体(磷酸多立醇)上。然后将其作为完整单元转移到序列Asn-X-Ser或Asn-X-Thr内的受体天冬酰胺(Asn)残基(其中X是除脯氨酸以外的任何氨基酸)。

Figure 7.26Synthesis of N-linked glycoproteins
The first step in glycosylation is the addition of an oligosaccharide consisting of 14 sugar residues to a growing polypeptide chain in the endoplasmic reticulum (ER). The oligosaccharide (which consists of two N-acetylglucosamine, nine mannose, and three glucose residues) is assembled on a lipid carrier (dolichol phosphate) in the ER membrane. It is then transferred as a unit to an acceptor asparagine residue of the polypeptide.
在进一步加工中,对常见的N-连接寡糖进行了修饰。当糖蛋白在内质网中时,三个葡萄糖残基和一个甘露糖被去除。寡糖在高尔基体中被进一步修饰,糖蛋白从内质网转移到高尔基体。这些修饰(将在第9章中讨论)包括当糖蛋白通过高尔基体的隔室运输时去除和添加碳水化合物残基(图7.27)。不同糖蛋白的N-连接寡糖被加工到不同程度,这取决于存在于不同细胞中的酶以及寡糖对催化其修饰的酶的可及性。在高尔基体中,具有难以接近的低聚糖的糖蛋白不会添加新的糖。这些糖蛋白中相对简单的低聚糖被称为高甘露糖低聚糖,因为它们含有高比例的甘露糖残基,类似于最初添加在内质网中的常见低聚糖。相反,具有可获得低聚糖的糖蛋白被更广泛地加工,从而形成各种复杂的低聚糖。
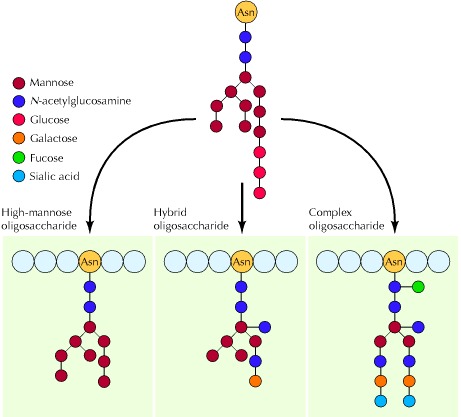
Figure 7.27Examples of N-linked oligosaccharides
Various oligosaccharides form from further modifications of the common 14-sugar unit initially added in the endoplasmic reticulum (see Figure 7.26). In high-mannose oligosaccharides, the glucose residues and some mannose residues are removed, but no other sugars are added. In the synthesis of complex oligosaccharides, more mannose residues are removed and other sugars are added. Hybrid oligosaccharides are intermediate between high-mannose and complex oligosaccharides. The structures shown are representative examples.
O-连接的低聚糖也被添加到高尔基体中。与N-连接的低聚糖不同,O-连接的低聚糖是通过一次添加一个糖形成的,通常只由几个残基组成(图7.28)。许多细胞质和核蛋白,包括各种转录因子,也通过添加单个O-连接的N-乙酰葡糖胺残基,在不同的酶系统催化下进行修饰。然而,碳水化合物在这些细胞质和细胞核糖蛋白功能中的作用尚不清楚。
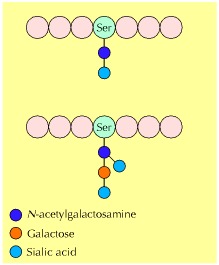
Figure 7.28Examples of O-linked oligosaccharides
O-linked oligosaccharides usually consist of only a few carbohydrate residues, which are added one sugar at a time.
五、脂质附着 Attachment of Lipids
真核细胞中的一些蛋白质通过脂质附着在多肽链上而被修饰。这种修饰经常将这些蛋白质靶向并锚定在质膜上,疏水性脂质能够与质膜相互作用(见图2.48)。三种常见的脂质添加类型——N-肉豆蔻酰化、异戊二烯化和棕榈酰化——在与质膜胞质面相关的真核蛋白质中很常见。第四种类型的修饰,糖脂的添加,在将一些细胞表面蛋白锚定到质膜的细胞外表面方面发挥着重要作用。
。。。。
参考资料
