【9.6.1.2】3UTR
在分子遗传学中,3‘端非翻译区( 3'-UTR ) 是紧跟在翻译终止密码子之后的信使 RNA (mRNA) 部分。3'-UTR 通常包含转录后影响基因表达的调节区。
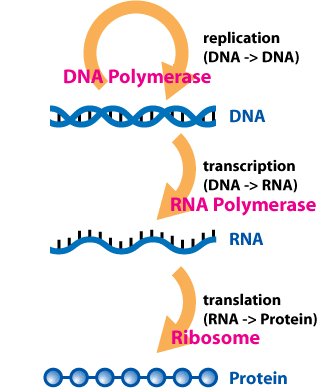
在基因表达过程中,一个 mRNA 分子从DNA序列中转录出来,然后被翻译成蛋白质。mRNA 分子的几个区域没有翻译成蛋白质,包括5' 帽、5' 非翻译区、3' 非翻译区和poly(A) 尾。3'-非翻译区内的调控区可以影响mRNA 的多聚腺苷酸化、翻译效率、定位和稳定性。[1] [2]
- 3'-UTR 包含调节蛋白和microRNA的结合位点(miRNA)。通过与 3'-UTR 内的特定位点结合,miRNA 可以通过抑制翻译或直接导致转录物降解来降低各种 mRNA 的基因表达。
- 3'-UTR 还具有与阻遏蛋白结合并抑制 mRNA 表达的 沉默区域。
- 许多 3'-UTR 还包含富含 AU 的元素(ARE)。蛋白质结合 ARE 以局部方式影响转录物的稳定性或衰减率或影响翻译起始。
- 此外,3'-UTR 包含序列 AAUAAA,该序列指导将数百个称为poly(A) 尾的腺嘌呤残基添加到 mRNA 转录物的末端。Poly(A) 结合蛋白 (PABP) 与该尾部结合,有助于调节 mRNA 翻译、稳定性和输出。例如,poly(A) 尾部结合的 PABP 与转录物 5' 端相关的蛋白质相互作用,导致促进翻译的 mRNA 环化。
- 3'-UTR 还可以包含吸引蛋白质以将 mRNA 与细胞骨架结合、将其运输到细胞核或从细胞核运输或执行其他类型定位的序列。除了 3'-UTR 内的序列外,该区域的物理特征,包括其长度和二级结构,也有助于翻译调控。这些不同的基因调控机制确保正确的基因在适当的时间在正确的细胞中表达。

一、物理特性
mRNA 的 3'-UTR 具有多种调节功能,受该区域的物理特性控制。其中一个特征是 3'-UTR 的长度,它在哺乳动物基因组中具有相当大的差异。mRNA 转录本的这个区域可以从 60个核苷酸到约 4000个核苷酸不等。 [3]人类 3'-UTR 的平均长度约为 800 个核苷酸,而 5'-UTR 的平均长度仅为约 200 个核苷酸。[4]3'-UTR 的长度很重要,因为较长的 3'-UTR 与较低水平的基因表达相关。对这种现象的一种可能解释是,较长的区域更有可能拥有更多具有抑制翻译能力的 miRNA 结合位点。除了长度之外,5' 和 3'-UTR 之间的核苷酸组成也有显着差异。温血脊椎动物 5'-UTR的平均G+C 百分比约为 60%,而 3'-UTR 仅为 45%。这很重要,因为在 5' 和 3'-UTR 的 G+C% 及其相应长度之间观察到了反相关。GC-poor 的 UTR 往往比位于富含 GC 的基因组区域的 UTR 长。[4]
- 3'-UTR 内的序列也具有降解或稳定 mRNA 转录物的能力。控制转录物稳定性的修饰允许在不改变翻译速率的情况下快速控制基因的表达。3'-UTR 中可以帮助破坏 mRNA 转录物稳定性的一组元素是富含 AU 的元素(AREs)。这些元件的大小范围为 50 到 150 个碱基对,通常包含多个拷贝的五核苷酸 AUUUA。早期研究表明,AREs 可以在序列上有所不同,并分为三个主要类别,它们在基序的数量和排列上有所不同。[1]
- 5' 和 3'-UTR 中存在的另一组元素是铁响应元素(IRE)。IRE 是 mRNA 非翻译区域内的茎环结构,编码参与细胞铁代谢的蛋白质。取决于特定蛋白质的结合和细胞内铁浓度,含有该元件的 mRNA 转录物要么被降解,要么被稳定。[3]
- RNA分子的茎环结构3'-UTR 还包含向转录本本身或向翻译产物发出信号的序列。例如,在 3'-UTR 内存在两种不同的多聚腺苷酸化信号,表明添加了多聚 (A) 尾。这些信号以约 250 个碱基对的确定长度启动 poly(A) 尾的合成。[1]使用的主要信号是核聚腺苷酸化信号 (PAS),其序列 AAUAAA 位于 3'-UTR 末端。[3]
然而,在早期发育过程中,细胞质多聚腺苷酸化可以代替发生并调节母体mRNA的翻译激活。控制这一过程的元素称为 CPE,它富含 AU,也位于 3'-UTR 中。CPE通常具有UUUUUUAU结构并且通常在核PAS的100个碱基对内。[3]由 3'-UTR 发出信号的另一个特定添加是在编码硒蛋白的 mRNA 的 UGA 密码子处掺入硒代半胱氨酸。通常 UGA 密码子编码停止翻译,但在这种情况下,称为硒代半胱氨酸插入序列 (SECIS)的保守茎环结构会导致硒代半胱氨酸的插入
二、在基因表达中的作用
3'-非翻译区通过影响 mRNA 的定位、稳定性、输出和翻译效率在基因表达中起着至关重要的作用。它包含参与基因表达的各种序列,包括 microRNA 反应元件 (MRE)、富含 AU 的元件 (ARE) 和 poly(A) 尾。此外,3'-UTR 的结构特征以及它对替代多聚腺苷酸化的使用在基因表达中起作用。
MicroRNA 反应元件
3'-UTR 通常包含 microRNA 反应元件 (MRE),它们是 miRNA 结合的序列。miRNA 是短的非编码 RNA 分子,能够与 mRNA 转录物结合并调节其表达。一种 miRNA 机制涉及 miRNA的 5' 种子序列与 mRNA 的 3'-UTR 内的 MRE 的部分碱基配对。这种结合会导致翻译抑制。
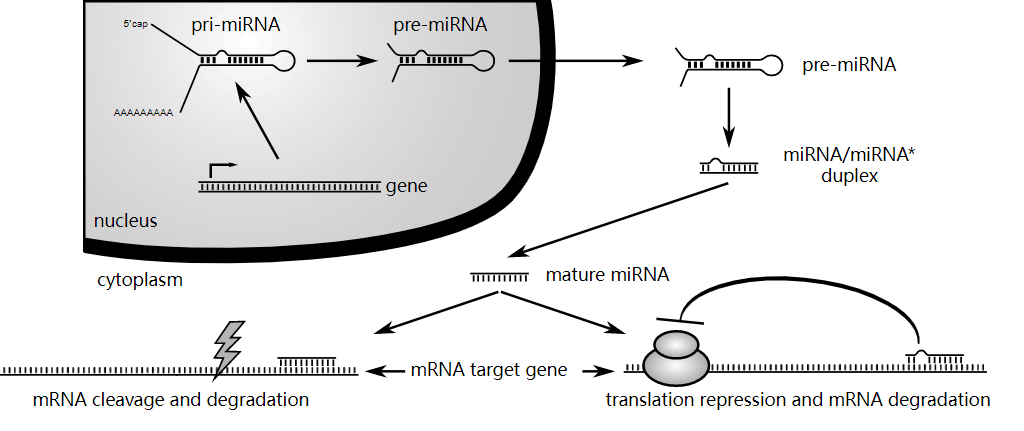
miRNA在基因调控中的作用
富含 AU 的元素
除了包含 MRE,3'-UTR 还经常包含富含 AU 的元素 (ARE),其长度为 50 至 150 bp,通常包含序列 AUUUA 的许多拷贝。ARE 结合蛋白 (ARE-BP) 以取决于组织类型、细胞类型、时间、细胞定位和环境的方式与富含 AU 的元素结合。针对不同的细胞内和细胞外信号,ARE-BPs可以促进mRNA衰变、影响mRNA稳定性或激活翻译。这种基因调控机制涉及细胞生长、细胞分化和对外部刺激的适应。因此,它作用于编码细胞因子、生长因子、肿瘤抑制因子、原癌基因、细胞周期蛋白、酶、转录因子、受体和膜蛋白。
Poly(A) 尾巴
聚 (A) 尾含有聚 (A) 结合蛋白 (PABP) 的结合位点。这些蛋白质与其他因素合作以影响 mRNA 的输出、稳定性、衰变和翻译。与 poly(A) 尾结合的 PABP 也可能与与 mRNA 5' 帽结合的蛋白质相互作用,例如翻译起始因子。这种相互作用导致转录物的环化,从而促进翻译起始。此外,它通过引起核糖体的回收来实现有效的翻译。[1] [2]虽然 poly(A) 尾的存在通常有助于触发翻译,但缺少或去除尾通常会导致外切核酸酶介导的 mRNA 降解。多聚腺苷酸化本身受转录本 3'-UTR 内的序列调控。这些序列包括细胞质多腺苷酸化元件 (CPE),它们是富含尿苷的序列,有助于多腺苷酸化的激活和抑制。CPE 结合蛋白 (CPEB) 与多种其他蛋白质结合到 CPE 上,以引发不同的反应。
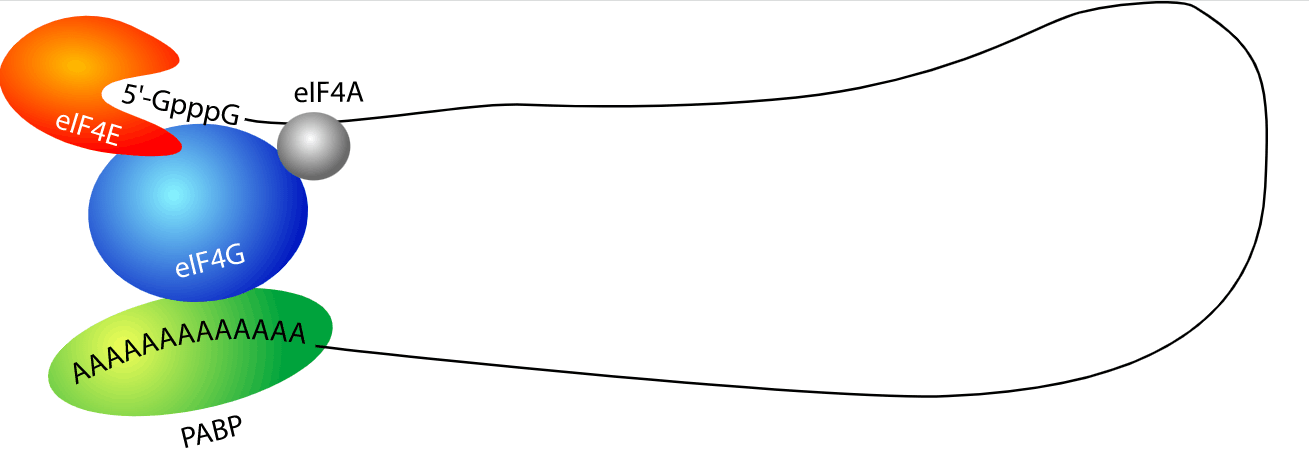
mRNA 转录物的环化由与 5' 帽和 poly(A) 尾相互作用的蛋白质介导
结构特征
虽然构成 3'-UTR 的序列对基因表达有很大贡献,但 3'-UTR 的结构特征也起着重要作用。一般来说,较长的 3'-UTR 对应于较低的表达率,因为它们通常包含更多参与抑制翻译的 miRNA 和蛋白质结合位点。[1] [2] [5] 人类转录本具有 3'-UTR,其平均长度是其他哺乳动物 3'-UTR 的两倍。这一趋势反映了人类基因调控所涉及的高度复杂性。除了长度,3'-非翻译区的二级结构也具有调节功能。蛋白质因子可以帮助或破坏该区域折叠成各种二级结构。最常见的结构是茎环,它为影响转录物表达的 RNA 结合蛋白和非编码 RNA 提供支架。[1]
替代多聚腺苷酸化
涉及 3'-UTR 结构的另一种机制称为替代多聚腺苷酸化 (APA),它导致 mRNA同工型仅在其 3'-UTR 上有所不同。这种机制对复杂的生物体特别有用,因为它提供了一种表达相同蛋白质但数量和位置不同的方法。它被大约一半的人类基因利用。APA 可能是由于存在多个多聚腺苷酸化位点或相互排斥的末端外显子。由于它可以影响蛋白质和 miRNA 结合位点的存在,APA 可以通过影响其稳定性、输出到细胞质和翻译效率来导致 mRNA 转录物的差异表达。[1] [5]
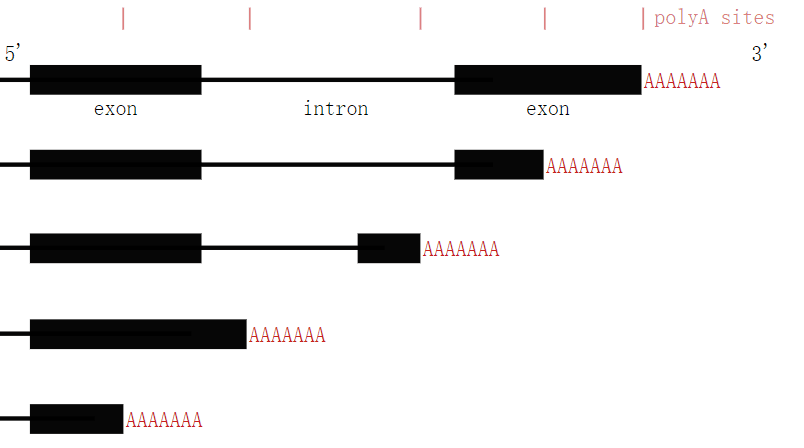
可选择的多聚腺苷酸化导致具有不同 3'-UTR 的转录本
三、研究方法
科学家们使用多种方法来研究 3' UTR 的复杂结构和功能。即使 mRNA 中的给定 3'-UTR 显示存在于组织中,也必须确定定位、功能半衰期、翻译效率和反式作用元件的影响,以了解 3'-UTR 的全部功能. [7]主要通过序列分析的计算方法表明,大约 5% 到 8% 的人类 3'-UTR 中存在 ARE,并且在多达 60% 或更多的人类 3'-UTR 中存在一个或多个 miRNA 靶标。软件可以一次快速比较数百万个序列,以发现基因组内各种 3' UTR 之间的相似性。实验方法已被用于定义与特定 RNA 结合蛋白相关的序列。具体而言,最近对测序和交联技术的改进使得能够在转录本中精确定位蛋白质结合位点。[8]诱导的位点特异性突变,例如影响终止密码子、多腺苷酸化信号或 3'-UTR 二级结构的突变,可以显示突变区域如何导致翻译失调和疾病。[9]这些类型的转录范围方法应该有助于我们了解 3'-UTR 中已知的顺式元件和反式调节因子。
四、疾病
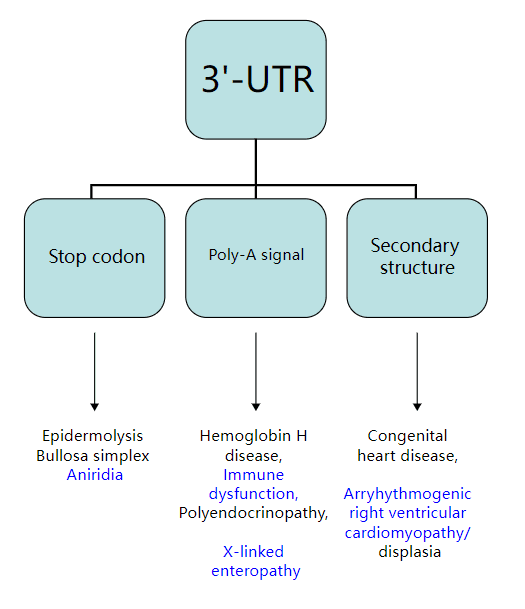
由 3'-UTR 内不同突变引起的疾病3'-UTR 突变可能非常重要,因为一种改变可能导致许多基因的表达改变。在转录上,突变可能只影响物理连接的等位基因和基因。然而,由于 3'-UTR 结合蛋白也在 mRNA 的加工和核输出中起作用,因此突变也会影响其他不相关的基因。[9]由于富含 AU 的区域的突变导致 ARE 结合蛋白 (AUBP) 的失调可导致疾病,包括肿瘤发生(癌症)、造血系统恶性肿瘤、白血病和发育迟缓/自闭症谱系障碍。[10] [11] [12] 3'-UTR 中三核苷酸 (CTG) 重复的扩展数量肌强直性肌营养不良蛋白激酶(DMPK)基因导致肌强直性营养不良。[7] fukutin 蛋白 3'-UTR 内串联重复序列的逆向转座 3 千碱基插入与 Fukuyama 型先天性肌营养不良症有关。[7] 3'-UTR 中的元素也与人类急性髓性白血病、α-地中海贫血、神经母细胞瘤、角化病、无虹膜、IPEX 综合征和先天性心脏缺陷有关。[9]确定的少数 UTR 介导的疾病仅暗示了尚未发现的无数联系。
五、未来发展
尽管我们目前了解 3'-UTR,但它们仍然是相对的谜。由于 mRNA 通常包含几个重叠的控制元件,因此通常很难指定每个 3'-UTR 元件的身份和功能,更不用说可能在这些位点结合的调节因子了。此外,每个 3'-UTR 包含许多替代的富含 AU 的元素和多聚腺苷酸化信号。这些顺式和反式作用元件,连同 miRNA,在单个 mRNA 中提供了几乎无限范围的控制可能性。[7]通过增加使用基于深度测序的核糖体分析的未来研究将揭示更多的监管细节以及新的控制元件和 AUBP。[1]此外,成绩单的最终命运在于它所涉及的信号转导通路,因此该领域的未来研究似乎很有希望。
参考资料
