【1.4.3】免疫应答--B细胞介导的体液免疫应答
与T细胞不同,B细胞的抗原受体BCR可以直接识别抗原分子表面的抗原表位,不需要MHC的抗原递呈作用。B细胞对抗原的免疫应答有两种情况:
- 胸腺非依赖性抗原(thymus-independent antigen, TI抗原),大多数为多糖类和脂类抗原,不需要T细胞的辅助,就可以直接激活B细胞产生IgM类抗体,主要由B1细胞介导,其抗原受体多样性有限,特异性差;
- 胸腺依赖性抗原(thymus-dependent antigen ,TD抗原),为蛋白类抗原,激活B细胞发生免疫应答需要T细胞的辅助,其时间经历Ig亲和力成熟、类别转换和记忆B细胞形成等过程。
一、B 细胞对 TD 抗原的免疫应答
1.1 B 细胞对 TD 抗原的识别
B 细胞通过 BCR 识别抗原
BCR 识别抗原在 B 细胞激活中有两个相互 关联的作用:
- 抗原与 mIg 的可变区特异结合,产生活化第一信号;
- 内化与其结合 的抗原,并进行加工处理,形成抗原肽-MHCII类分子复合物,提呈给 Th 细胞识 别,Th 活化后反过来提供 B 细胞活化的第二信号。
前一个作用对于后一个作用 是必须的,抗原只有与 BCR 结合后,才能被 B 细胞内化。与 TCR 不同,BCR 识 别抗原有如下特点:
- BCR 不仅能识别蛋白质,还能识别多肽、核酸、多糖、脂 类和小分子化合物;
- BCR 可特异性识别完整抗原的天然构象;
- BCR 识别抗 原无需 APC 加工处理,也没有 MHC 限制性。
B 细胞的活化信号
B 细胞活化也需要双信号:
- BCR 与抗原特异性结合, 启动 B 细胞活化的第一信号
- 由共刺激分子提供的第二信号使 B 细胞完全活化。 活化后的信号转导途径与 T 细胞类似。
(1)B 细胞活化的第一信号:B 细胞活化的第一信号经由 BCR-CD79a/b(即 Igα/Igβ) 和 CD19/CD21/CD81 共同传递。抗原与 BCR 结合启动的活化信号, 需要 CD79a/b 将之传入胞内。与 CD3 类似,CD79a/b 胞浆区也存在 ITAM。ITAM 磷酸化后,启动激酶活化的级联反应,最终通过激活转录因子,调控细胞增殖及 活化相关基因,使 B 细胞初步活化。
(2)BCR 共受体的作用:成熟 B 细胞表面 CD19/CD21/CD81 以非共价键 组成 BCR 共受体复合物。CD21 胞内区无酪氨酸残基,故不能传导信号,但能识 别结合于抗原的补体成分 C3d,与 CD19 交联,向胞内传递信号。CD19 的胞浆 区有多个保守的酪氨酸残基,能募集含有 SH2 结构域的酪氨酸激酶 Lyn、Fyn 等。 CD81 分子为 4 次跨膜分子,其主要作用可能是联结 CD19 和 CD21,稳定 CD19/CD21/CD81 复合物。共受体传导的信号加强了由 BCR 复合物传导的信号, 明显降低了抗原激活 B 细胞的阈值,从而大大提高了 B 细胞对抗原刺激的敏感 性。
(3)B 细胞活化的第二信号:B 细胞的完全活化,需要由 Th 细胞与 B 细胞 表面多对共刺激分子相互作用提供的共刺激信号(第二信号),其中最重要的是 CD40/CD40L。CD40 主要表达在 B 细胞以及单核细胞和 DC 表面;CD40L 主要 表达在活化的 CD4+T 细胞和肥大细胞的表面。静息 T 细胞不表达 CD40L,活化T 细胞迅速表达 CD40L,CD40L 与 B 细胞表面组成性表达的 CD40 相互作用, 向 B 细胞传递活化的第二信号。
(4)细胞因子的作用:活化 B 细胞表达多种细胞因子受体,在活化 T 细胞 分泌的细胞因子作用下大量增殖。细胞因子诱导的 B 细胞增殖是 B 细胞形成生 发中心和继续分化的基础。
1.2 T、B 细胞相互作用与 B 细胞免疫应答
B 细胞对 TD 抗原的应答需要 T 细胞辅助,表现为:
- 活化 T 细胞表面的共 刺激分子可以提供 B 细胞活化的第二信号;
- T 细胞分泌的细胞因子促进 B 细 胞活化、增殖、分化。
T、B 细胞间的作用是相互的。
- 一方面,B 细胞可以作为 APC 加工、提呈抗 原肽-MHCII类分子复合物给 T 细胞识别,活化 T 细胞;
- 另一方面,活化 T 细胞 提供 B 细胞活化的第二信号,并分泌 IL-4 等细胞因子协助 B 细胞的进一步分化。
Th 细胞在生发中心暗区的形成、B 细胞克隆性扩增和 B 细胞分化成生发中心细 胞、抗体类别转换以及记忆性 B 细胞的生成中均起重要作用。
体内抗原特异性 B 细胞与抗原特异性 T 细胞相遇的几率非常小,仅为 1/10^8~ 1/10^12。而在外周淋巴器官中,T、B 细胞定位于不同的部位,这就产生了抗原特 异 T 细胞如何辅助抗原特异 B 细胞的问题。这可能与迁移中的抗原提呈细胞对 抗原的捕捉相关。抗原一旦进入机体,会被专职 APC,特别是树突状细胞捕捉和 加工,后者从组织迁移到局部淋巴结的 T 细胞区。经抗原刺激的初始 T 细胞在 再循环中连续不断流过这些细胞旁边时,那些有着能识别抗原中 T 细胞表位的 T 细胞就被 APC“捕捉”。被“捕捉”的 T 细胞的 TCR 与 APC 加工提呈的抗原肽 -MHC 分子相结合,T 细胞被激活。血循环中的 B 细胞也穿越高内皮小静脉进入 T 细胞区,表达该抗原特异性 BCR 的 B 细胞也在该区被“捕捉”。这样,抗原特 异的 B 细胞就能与抗原特异的 Th 细胞在 T 细胞区这一特定的部位相遇,B 细胞 就能在 Th 细胞辅助下被激活,再进入淋巴滤泡。进入淋巴滤泡的 B 细胞分裂增 殖,形成生发中心(germinal center)。生发中心里的 B 细胞会多次离开生发中心, 再回到生发中心。Th 细胞也能进入生发中心,也在生发中心给 B 细胞予辅助。 部分激活的 B 细胞分化成浆细胞,产生抗体。部分激活的 B 细胞分化成记忆细 胞。上述 T 细胞对 B 细胞体液免疫应答的辅助均为特异性辅助,即 T 细胞辅助 的 B 细胞所识别的与 T 细胞识别的为同一抗原。
1.3 B 细胞的增殖和终末分化
初始 B 细胞经双信号刺激而完全活化,具备了增殖和继续分化的能力。在 Th 细胞产生的细胞因子辅助下,活化 B 细胞增殖形成生发中心,并经历体细胞 高频突变、Ig 亲和力成熟、Ig 类别转换,分化为浆细胞或记忆 B 细胞,发挥免 疫效应。在 B 细胞活化、增殖与分化过程中,均需 Th 细胞的辅助。
B 细胞在生发中心的分化成熟
在外周免疫器官 T 细胞区激活的部分 B 细胞进入初级淋巴滤泡,分裂增殖,形成生发中心。生发中心是 B 细胞增殖分化 的主要场所,在抗原刺激后一周左右形成。其中的 B 细胞 6 ~8 小时分裂一次。 这些分裂增殖的 B 细胞称中心母细胞(centroblast),有着 B 细胞的典型形态特 征,分裂能力极强。不发生分裂增殖的 B 细胞被推向外侧,形成冠状带(mantle zone)。随着生发中心细胞增加,生发中心可分为两个区域:一个是暗区(dark zone), 分裂增殖的中心母细胞在此紧密集聚,滤泡树突状细胞(FDC)很少;另一个为 明区(light zone),中心细胞在此聚集不甚紧密,但与众多的 FDC 接触(图 8- 4)。生发中心中绝大多数 B 细胞发生凋亡。部分 B 细胞在抗原刺激和 T 细胞辅 助下继续分化发育,并可发生体细胞高频突变、Ig 亲和力成熟、Ig 类别转换及记 忆 B 细胞形成等变化。
体细胞高频突变和 Ig 亲和力成熟
中心母细胞重链和轻链 V 区基因的 体细胞突变率极高,每次细胞分裂有 50% BCR 突变,是形成抗体多样性的主要 机制之一。由于体细胞高频突变,导致 BCR 的特异性或亲和力发生改变,从而 形成多样性 B 细胞克隆,只有那些表达高亲和力抗原受体的 B 细胞,才能有效 地结合抗原,并在抗原特异性 Th 细胞辅助下增殖,产生高亲和力的抗体,此为 抗体亲和力成熟(affinity maturation)。
Ig 类别转换
B 细胞在 IgV 区基因重排后,其子代细胞 Ig 重链 V 区基因 保持不变,但 C 区基因则会发生重排。在免疫应答中首先分泌 IgM,但随后可表 达并产生 IgG、IgA 或 IgE,而其重链 V 区不发生改变。这种可变区相同而 Ig 类 别发生变化的过程称为 Ig 类别转换(class switching)或同种型转换(isotype switch)。Ig 类别转换在抗原诱导下发生,并接受 T 细胞分泌的细胞因子调节。 Ig 类别转换是机体产生不同类别抗体的基础。
浆细胞的形成
浆细胞又称抗体形成细胞(antibodyformingcell,AFC), 是 B 细胞分化的终末细胞,浆细胞胞质中除了少量线粒体,几乎全部为粗面内质网,能合成和分泌特异性抗体,同时表面的 BCR 表达减少。与初始 B 细胞不同, 浆细胞的主要特点是能够分泌大量抗体,而不能与抗原起反应,也失去了与 Th 细胞相互作用的能力,因为浆细胞表面不再表达 BCR 和 MHCII类分子。生发中 心产生的浆细胞大部分迁入骨髓,并在较长时间内持续产生抗体。
记忆性 B 细胞的产生
生发中心中存活下来的 B 细胞,或分化发育成浆细胞, 或成为记忆细胞离开生发中心。记忆 B 细胞(memory B cell,Bm)不产生 Ig, 但再次与同一抗原相遇时可迅速活化,产生大量抗原特异性 Ig。记忆 B 细胞表 达 CD27,并较初始 B 细胞表达较高水平的 CD44。一般认为记忆细胞为长寿细 胞,尚不清楚是什么因素维持记忆细胞的存活。有人认为抗原可能以 IC 状态持 续存在于生发中心中 FDC 的树突表面,它能“不断”刺激“经过”生发中心的 记忆 B 细胞,给后者存活信号。
二、B 细胞对 TI 抗原的免疫应答
非胸腺依赖性抗原(TI-Ag),如某些细菌多糖、多聚蛋白质及脂多糖等,能 直接刺激初始 B 细胞活化,而无需抗原特异性 T 细胞的辅助。在正常个体,TI 抗原可诱导抗体产生,而不引起 T 细胞应答。TI 抗原可分成两类,即 TI-1 和 TI- 2,它们以不同机制激活 B 细胞。
2.1 B 细胞对 TI-1 抗原的免疫应答
TI-1 抗原常被称为 B 细胞丝裂原,可以诱导成熟和未成熟 B 细胞活化,产 生低亲和力的 IgM。高浓度 TI-1 抗原经丝裂原受体与 B 细胞结合,多克隆地诱 导 B 细胞增殖和分化;低浓度抗原则能激活抗原特异性 B 细胞。在机体感染病 原体时,可能 TI-1 抗原的浓度很低,因此只有抗原特异的 B 细胞才能被激活, 并产生抗该抗原的抗体。该应答在抗某些胞外病原体感染中发挥重要作用,因其 无需 Th 细胞预先致敏与克隆性扩增,故比对胸腺依赖性抗原的应答发生为早。 但 TI-1 抗原单独不足以诱导 Ig 类别转换、Ig 亲和力成熟及记忆 B 细胞形成,这 些均需特异 T 细胞辅助。
2.2 B 细胞对 TI-2 抗原的免疫应答
TI-2 抗原为细菌胞壁与荚膜多糖,有高度重复的结构。TI-2 抗原只能激活成 熟 B 细胞,婴幼儿 B 细胞未发育成熟,故不能有效产生抗多糖抗原的抗体。对 TI-2 抗原发生应答的主要是 B1 细胞。TI-2 抗原通过其高度重复的抗原表位使 B 细胞的 mIg 发生广泛的交联而被激活,但 mIg 过度交联会使成熟 B 细胞变成无 应答性。因此,TI-2 抗原表位的密度在 TI-2 抗原激活 B 细胞中可能起决定作用。 密度太低,mIg 交联的程度不足以激活细胞;密度太高,会使细胞变为无应答性。
B 细胞对 TI-2 抗原的应答具有重要的生理意义。大多数胞外菌有胞壁多糖, 能抵抗吞噬细胞的吞噬消化,这使它们能逃脱吞噬细胞的直接吞噬杀伤。B 细胞 针对此类 TI-2 抗原产生的抗体,可发挥调理作用,促进吞噬细胞对病原体的吞 噬,并且有利于巨噬细胞将抗原提呈给特异性 T 细胞。TI-2 抗原可以直接激活 B1 细胞,T 细胞分泌的细胞因子可明显增强此类 B 细胞的免疫应答,并发生抗 体类别转换,可产生 IgM 及 IgG。T 细胞在 TI-2 抗原应答中的作用机制尚不十 分清楚。
三、体液免疫应答的一般规律
在抗原诱导下,浆细胞产生的抗体经淋巴液和血液流向全身,血流中抗体的 浓度随应答时间的持续而增高。
- 在初次接受特定抗原刺激时,机体发生的应答称 初次应答(primary response);
- 初次应答中所形成的记忆细胞再次接受相同抗原 刺激,产生迅速、高效、持久的应答,称为再次应答(secondary response),或称 回忆应答(anamnestic response)。
1.1 初次应答
初次应答产生的抗体数量少、亲和力低,产生过程可以分为如下四个阶段。
- 潜伏期(lagphase) 接受抗原刺激后到血清中能测到特异抗体之前的阶 段。此期的长短由抗原的性质、抗原进入机体的途径、所用佐剂类型及宿主情况 所决定,可短至数小时,也可长至数周。
- 对数期(log phase) 此期抗体量呈指数增加,取决于抗原剂量和抗原性 质等因素。
- 平台期(plateau phase) 此期血清中抗体浓度基本维持在一个相当稳定 的较高水平。到达平台期所需的时间及平台的高度及维持时间,依抗原不同而异。 有的平台期很短,有的可长至数周。
- 下降期(decline phase) 由于抗体被降解或与抗原结合而被清除,血清 中抗体浓度慢慢下降,此期可持续几天或几周。
1.2 再次应答
当再次接受相同抗原刺激,机体可发生再次免疫应答。它与初次应答的不同 之处为:
- 潜伏期短,大约为初次应答潜伏期时间的一半;
- 血清抗体浓度增加 快,到达平台期快,平台高;
- 维持时间长,下降期持久,因为机体会长时间合 成抗体;
- 用较少量抗原刺激即可诱发再次应答;
- 再次应答中产生的抗体主要 为高亲和力的 IgG,而初次应答中主要产生低亲和力的 IgM。
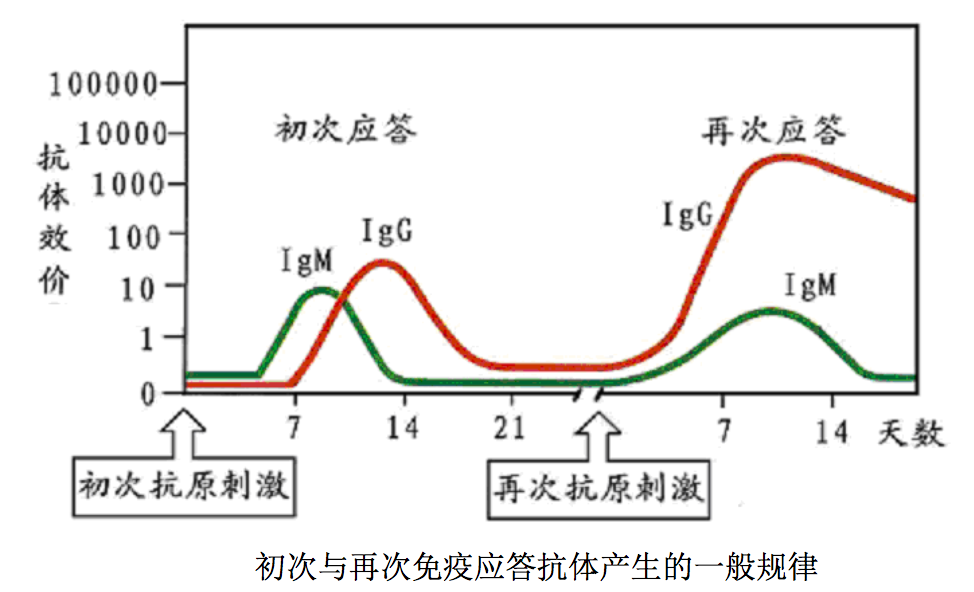
再次应答的强弱取决于抗原的强弱与两次抗原注射的间隔长短。间隔短则应 答弱,因为初次应答后存留的抗体可与注入的抗原结合,形成抗原-抗体复合物 而被迅速清除。间隔太长,反应也弱,因为记忆细胞尽管长命,但并非永生。再 次应答的能力可持续存在数个月或数年,故机体一旦被感染后可持续相当时间不 再感染相同病原体。
四、体液免疫应答的效应
抗体是浆细胞合成分泌的特异性免疫分子,主要通过以下作用方 式发挥抗感染等免疫作用:
- 中和作用:抗体与相应细菌毒素或病 原体特异性结合后,可阻止细菌毒素或病原体对易感细胞的侵入或感 染。
- 调理作用:IgG 类抗体与相应细菌等颗粒性抗原特异性结合 后,通过其 Fc 段与吞噬细胞表面 IgGFcR 结合,可增强吞噬细胞对细 菌等抗原性异物的吞噬杀伤或清除作用。
- 激活补体产生溶菌效应: 病原体特异性 IgG/IgM 类抗体与相应病原体结合后,可激活补体经典 途径产生 C5b6789 攻膜复合物使病原体溶解破坏。
- 抗体依赖细胞 介导的细胞毒作用(ADCC 作用):IgG 类抗体与病毒感染/肿瘤靶细胞 表面相应抗原表位特异性结合后,通过其 Fc 段与 NK 细胞/吞噬细胞 表面 IgGFcR 结合,可增强 NK 细胞/吞噬细胞对上述靶细胞的杀伤破 坏作用。
参考资料
- 《医学免疫学》课件 新乡医学院 宋向凤、张国俊、徐春阳、孙爱平、孙书明、赵铁锁等老师
