【2.1】免疫效应分子--免疫球蛋白抗体(Antibody,Ab)
一、抗体与免疫球蛋白的区别
- 抗体(Antibody,Ab):由浆细胞响应抗原产生的球蛋白并特异性结合特定的Ag。Ab通常根据它们与免疫Ag的特异性结合来定义
- 免疫球蛋白(Immunoglobulins ,Ig):Igs是球蛋白,其在化学结构中起到Ab或类似于Ab的作用。
并非所有Ig都具有Ab的功能,例如:多发性骨髓瘤患者的Ig水平非常高,但不是Ab
分泌Ig(Secreted Ig ,sIg)介导的体液免疫 膜Ig(Membrane Ig ,mIg):BCR B细胞受体
抗体(antibody,Ab)是B淋巴细胞接受抗原刺激后增殖分化为浆细胞,由浆细胞合成、分泌能与相应抗原特异性结合的免疫球蛋白(immunoglobulin,lg)。
抗体主要存在于血清中,也存在于各种体液中。

二、Ig的基本结构及多样性
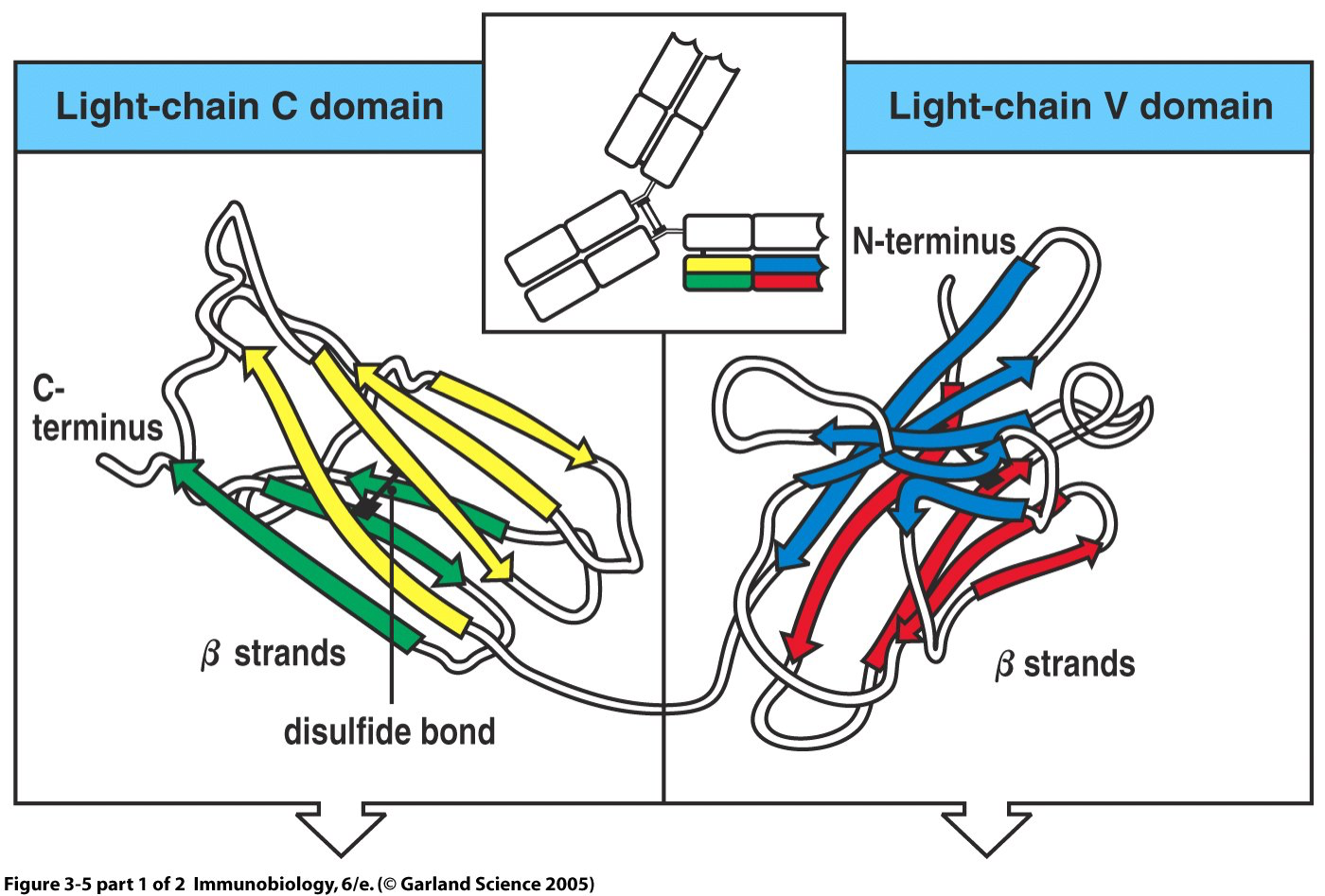
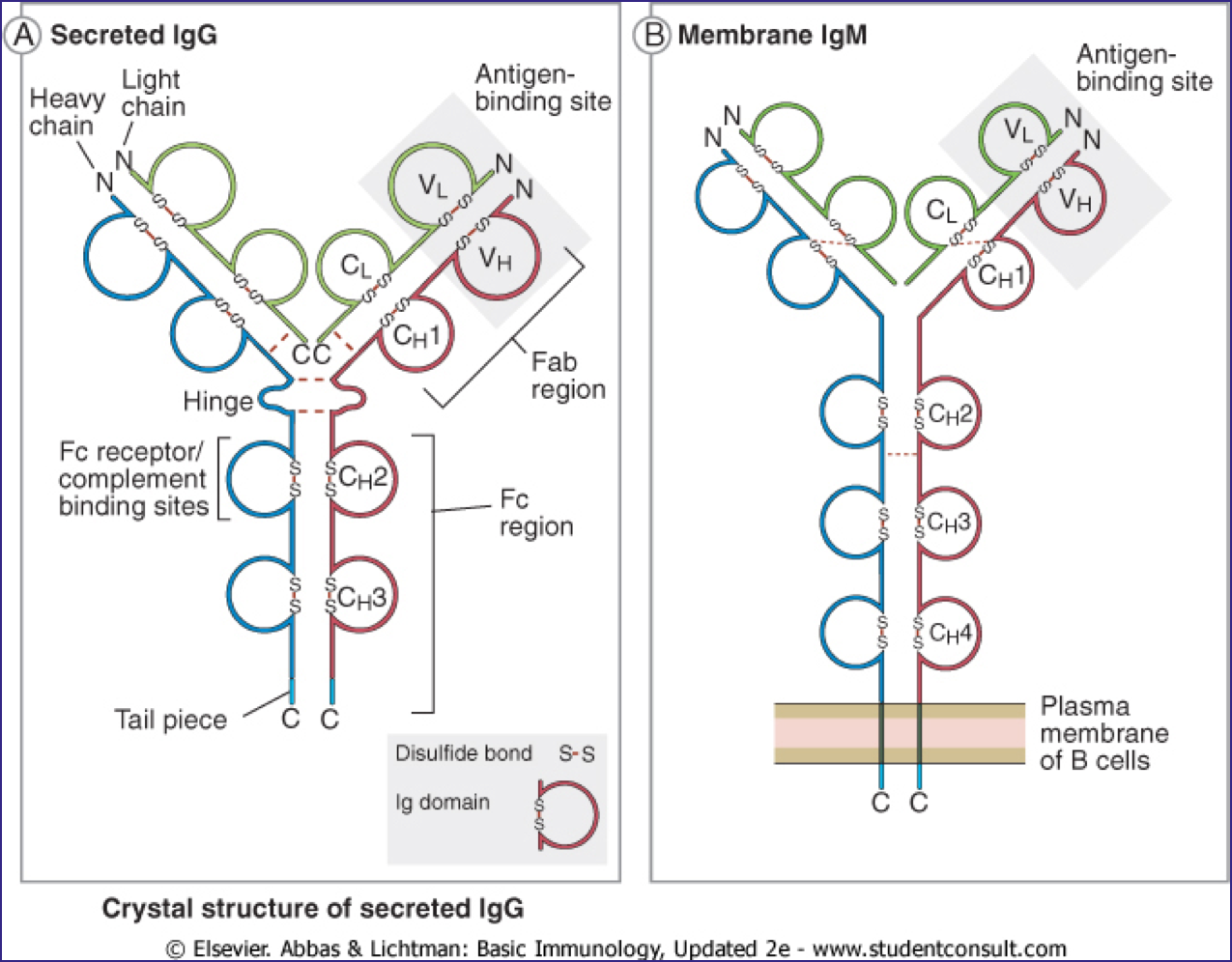
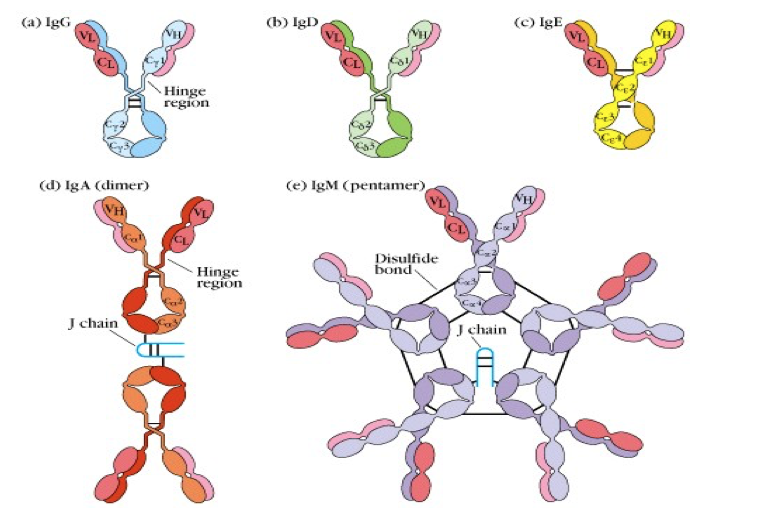
Ig分子的基本结构是由两条相同的重链和两条相同的轻链通过二硫键连接而成的四肽链结构,称为Ig的单体。
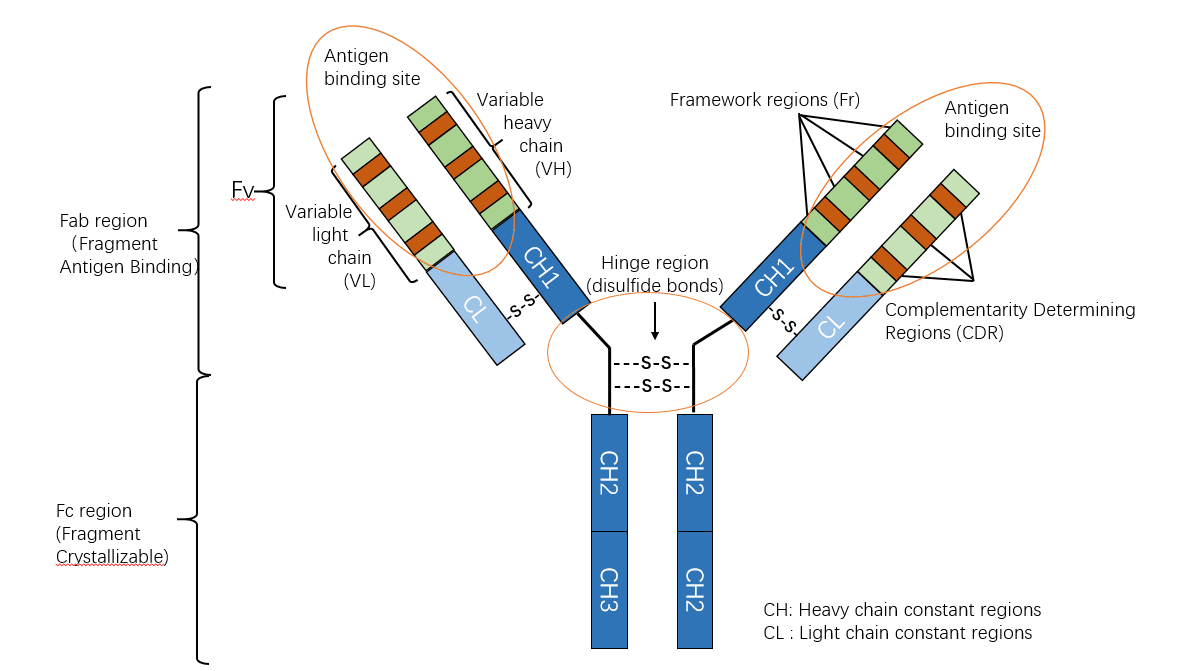
抗体的基本结构是由两条较长的多肽链和两条较短的多肽链通过二硫键连 接呈“Y”形构成。每条肽链分别由 2~5 个约含 110 个氨基酸,序列相似但功能 不同的结构域(又称功能区)组成。结构域的二级结构是由几股多肽链折叠形成 的两个反向平行的 β 片层经一个链内二硫键连接稳定的“β 桶状”结构。
2.1 重链(heavy chain, H链)
- Ig的重链由450~550个氨基酸残基组成,分子量约为50~75KD。
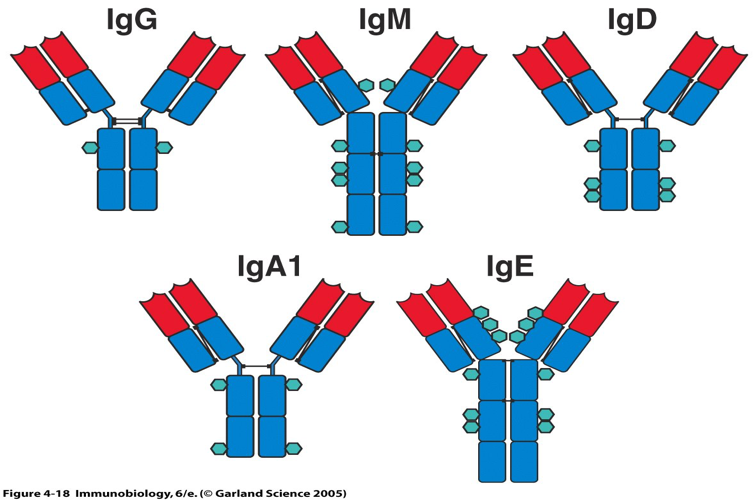
- 根据重链免疫原性不同,可将重链分为 μ ,γ, α ,δ,ε 五类(class),与轻链结合为 IgM, IgG, IgA, IgD和IgE。某些又可亚类。γ分为四个亚类(γ 1-4),α 分为2个亚类(α 1-2)。不同类的抗体 分子具有不同的特征及功能,如链内二硫键的数目和位置、结构域的数目、连接 寡糖的数量以及铰链区的长度等均不完全相同。即使是同一类抗体,铰链区氨基 酸组成和重链二硫键的数目、位置也不同,据此可将其分为不同的亚类。如人 IgA 可分为 IgA1 和 IgA2;IgG 可分为 IgG1~IgG4; IgM、IgE 和 IgD 尚未发现有亚 类。
- 重链的大小和组成不同; α和γ含有约450个氨基酸,而μ和ε含有约550个氨基酸。
- 每条重链具有两个区域,恒定区和可变区。恒定区在相同同种型的所有抗体中是相同的,但在不同同种型的抗体中不同。重链γ,α和δ具有由三个串联Ig结构域和铰链区组成的恒定区,用于增加柔性,重链μ和ε具有由四个免疫球蛋白结构域组成的恒定区。重链的可变区根据产生它的B细胞而不同,但对于由单个B细胞或B细胞克隆产生的所有抗体是相同的。每条重链的可变区长约110个氨基酸,由单个Ig结构域组成。
2.2 轻链(light chain, L链)
- Ig的轻链约含214个氨基酸,约为重链的1/2,分子量约为25 KD。
- 在哺乳动物中,轻链免疫原性的不同,分为κ、λ两型。
- 同一个体可存在分别带有为κ和λ链的Ig分子,同一天然Ig分子中不可能同时有κ和λ链,两条轻链的型号必然相同。
- 五类抗体中每 类抗体的轻链中都可以有 κ 链或 λ 链,两型轻链的功能无差异。不同种属生物体 内两型轻链的比例不同,正常人血清免疫球蛋白 κ:λ 约为 2:1,而在小鼠则为 20:1。 κ:λ 比例的异常可能反映免疫系统的异常,例如人类人类免疫球蛋白 λ 链过多, 提示可能有产生 λ 链的 B 细胞肿瘤。由于 λ 链恒定区个别氨基酸之间存在一定 差异,又可将 λ 链分为四个亚型:λ1、λ2、λ3 和 λ4。
- 轻链具有两个连续的域:一个恒定域和一个可变域。其他类型的轻链,例如iota(ι)链,存在于较低的脊椎动物中,如Chondrichthyes和Teleostei。
人体中的轻链
人类有两种类型的轻链:
- kappa(κ)链,由染色体2上的免疫球蛋白κ基因座(IGK)编码
- lambda(λ)链,由22号染色体上的免疫球蛋白λ基因座(IGL)编码
抗体由B淋巴细胞产生,每个B淋巴细胞仅表达一类轻链。 一旦设定,轻链类在B淋巴细胞的寿命期间保持固定。 在健康个体中,总血清与λ的比率在血清中大约为2:1(测量完整的完整抗体)或如果测量游离轻链则为1:1.5,具有指示肿瘤的高度不同比率。
根据一种新的多克隆游离轻链试验,kappa与λ的确切正常比例范围为0.26至1.65。κ和λ链都可以按比例增加,保持正常比例。 这通常表示除血细胞恶液质之外的其他事物,例如肾病。
其他动物中
四足动物中的免疫球蛋白轻链基因可分为三个不同的组:κ(κ),λ(λ)和西格玛(σ)。 κ,λ和σ同种型的分歧先于四足动物的辐射。两栖谱系进化后和爬行动物谱系出现之前,σ同种型丢失了。
其他类型的轻链可以在低等脊椎动物中找到,例如Chondrichthyes和Teleostei的Ig-Light-Iota链。
骆驼科动物在哺乳动物中是独一无二的,因为它们还具有完整功能的抗体,其具有两条重链,但缺少通常与每条重链配对的轻链。
作为其适应性免疫系统的一部分,鲨鱼还具有称为IgNAR(免疫球蛋白新抗原受体)的功能性重链同源二聚体抗体样分子。据信IgNAR从未有过相关的轻链,这与骆驼科动物中仅有重链抗体可能通过进化失去其轻链伴侣的理解相反。[6] [7]
2.3 可变区 Fv (Variable region )
氨基酸排列顺序在不同抗体分子间变化较大称为可变区(variable region ,V区)
可变区Ig 近氨基酸轻链的1/2和重链的1/4(γ, α ,δ)或(μ 、ε )。
Fv : 为VL和VH
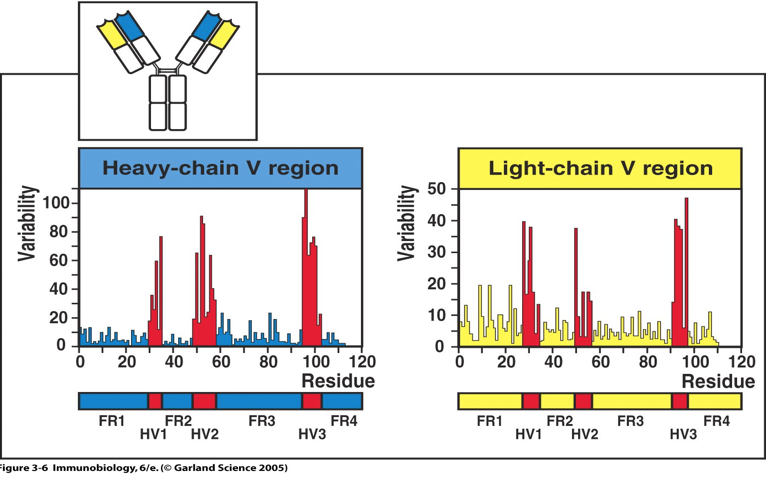
2.3.1 高变区(hypervariable region,HVR )
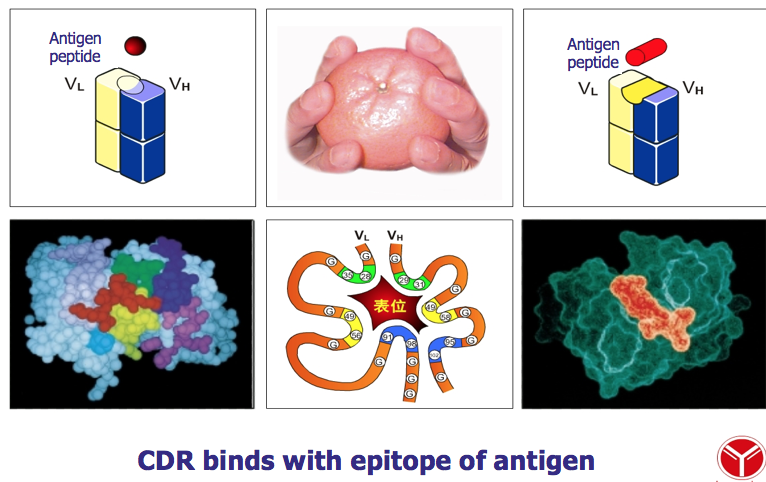
V 区中氨基酸的种类和排列顺序千变万化,故可形成种类极 多且具有不同特异性的抗体。重链 V 区(VH)和轻链 V 区(VL)各有 3 个区域 的氨基酸组成和排列顺序高度可变,称为高变区(hypervariable region,HVR);因 该区域形成与抗原表位互补的空间构象,又被称为互补决定区(complementarity determining region,CDR),分别用 CDR1(HVR1)、CDR2(HVR2)和 CDR3(HVR3) 表示,一般 CDR3 变化程度更高。VH 和 VL 共 6 个 CDR 共同组成 Ab 的抗原结 合部位(antigen-binding site),决定着抗体结合抗原的特异性,负责识别及结合抗 原,并发挥免疫效应。
CDR (complementarity-determining region): 因为这些序列形成与结合抗原的三维结构互补的表面,所以HVR也称为CDR
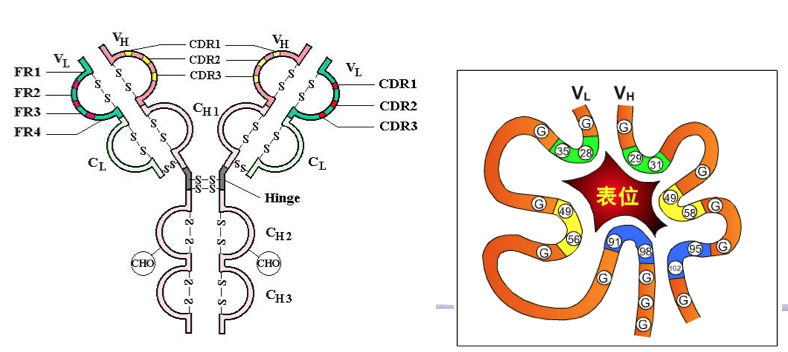
2.3.2 FR: 排除CDR区域的VL 或VH
V 区中的其他氨基酸残基称为骨架区或构架区(framework region,FR)。VL 或 VL 各有 4 个骨架区,分别用 FR1、FR2、FR3、FR4 表示, 大约占整个 V 区 75%,该区域的氨基酸组成和排列顺序变化不大。构架区的功能 是支持变变区并维持 V 区三维结构的稳定性。
2.4 F(ab) and Fc regions (Constant region ,恒定区)
重链和轻链的 C 区分别称为 CH 和 CL。不同型(κ 或 λ)抗体其 CL 的长度基本一致,但不同类抗体 CH的长度不一,其结构域数目也不同,如 IgG、IgA 和 IgD 重链 C 区有 CH1、CH2 和 CH3 三个结构域,IgM 和 IgE 重链 C 区有 CH1、CH2、CH3 和CH4四个结构域。同一种属的个体,所产生针对不同 抗原的同一类别抗体,尽管其 V 区各异,但其 C 区氨基酸的组成和排列顺序比较恒定,其免疫原性相同。例如,针对不同抗原的人 IgG 抗体,它们的 V 区不 同,所以只能与相应的抗原发生特异性结合,但 C 区是相同的,均含 γ 链,因 此抗人 IgG 抗体均能与之结合。
抗体的Y形可以分为三个部分:两个F(ab)区和Fc区。
- F(ab)区含有与同源抗原结合的可变结构域。
- Ig分子近C端,轻链的1/2和重链的3/4(或4/5)区域中氨基酸的数量、种类、排列,排列顺序及含糖量均较为恒定,称为恒定区(constant region,C区)。Fc片段为淋巴细胞表面上的内源性Fc受体提供结合位点,并且也是第二抗体结合的位点。此外,染料和酶可以与抗体Fc部分上的抗体共价连接,用于实验可视化。
- 重链的轻链的恒定区分别称为CH和CL。Ig的C区与抗体的生物学功能相关。
这三个区域可以通过蛋白水解酶胃蛋白酶切割成两个F(ab)和一个Fc片段。抗体片段在某些免疫化学技术中具有明显的优势。片段化IgG抗体有时是有用的,因为F(ab)片段(1)不会沉淀抗原,并且(2)由于缺乏Fc区而不会在活体研究中被免疫细胞结合。通常,由于它们较小的尺寸和缺乏交联(由于Fc区的丧失),F(ab)片段被放射性标记用于功能研究。 Fc片段通常在免疫组织化学染色中用作Fc受体阻断剂。
2.6 铰链区 (Hinge Region)
- 在CH1和CH2之间,即重链的链间二硫键连接处附近,有一个可转动的铰链区(hinge region)。
- 含较多的脯氨酸(proline )残基,富于柔性,可赋予较大的自由活动度。该区由于是一段较伸展的肽链,对蛋白酶敏感,易被木瓜蛋白酶、胃蛋白酶水解。
- IgM和IgE没有铰链区
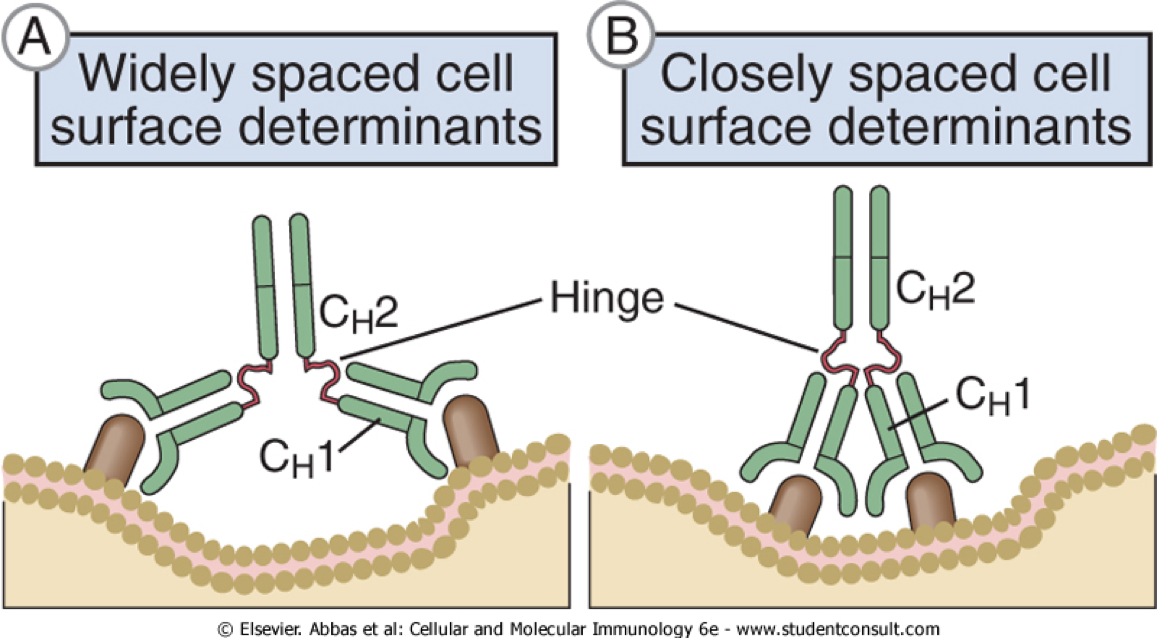

其功能包括:
- 当 VL、VH 与抗原结合时,铰链区伸展弯曲,使抗体分子上两 个抗原结合点更好地与相应的两个抗原表位互补;
- 当抗体与抗原结合时,铰链 区可使 Ig 分子的构型发生改变从“T”到“Y”,从而使位于 CH2 功能区的补体 结合点得以暴露,从而为补体经典途径激活提供条件。
2.7 Domains of immunoglobulin
Ig分子的3D图像显示它不是如图所示的笔直。 它被折叠成球状区域,每个区域包含链内二硫键。 这些区域称为域(domains)。
- Light Chain Domains:
- VL and CL
- Heavy Chain Domains
- VH, CH1, CH2, CH3 (IgA, IgD,IgG )
- CH4 in IgE and IgM
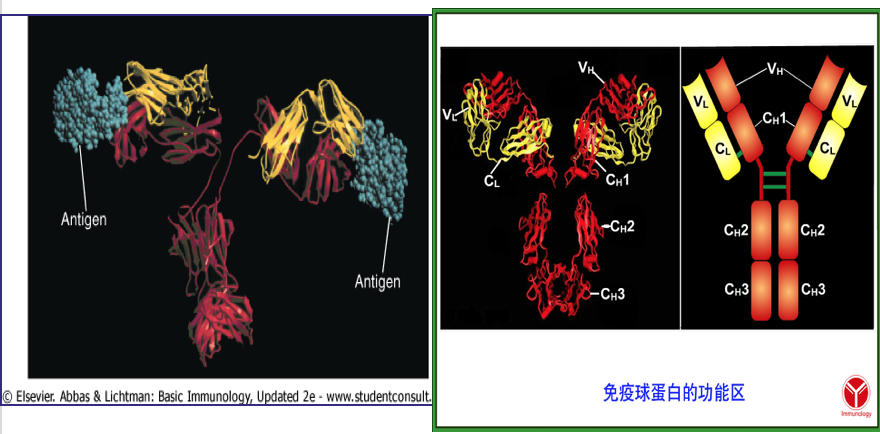
- VH和VL: 抗原结合部位
- CH1-3和CL: Ig 遗传标志
- CH2(IgG)、 CH3(IgM): 补体结合部位
- CH2-CH3(IgG):结合并通过胎盘
- CH3(IgG) CH4 (IgE):FcR结合部位
2.8 抗体的辅助成分
2.8.1 Joining chain
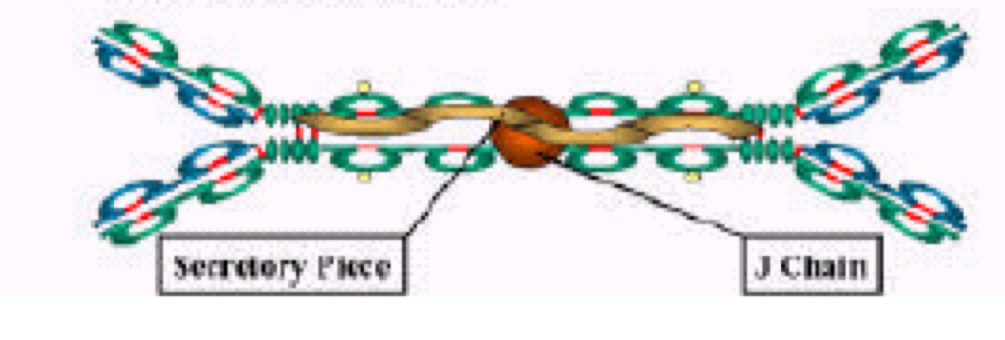
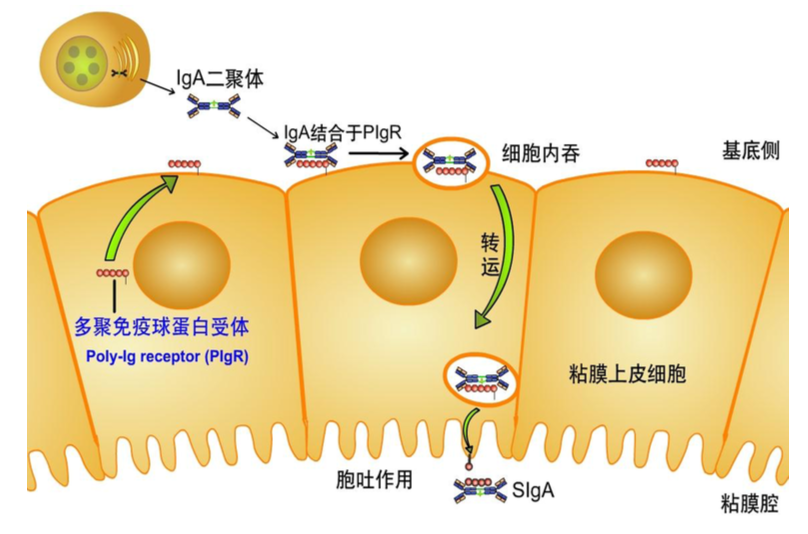
J 链(joining chain)是由 124 个氨基酸组成、富含半胱氨酸的酸性糖蛋白,分 子量约 15kD,由浆细胞合成;主要功能是将单体抗体分子连接为二聚体或多聚 体。2 个 IgA 单体由 J 链连接形成二聚体,5 个 IgM 单体由二硫键相互连接,并 通过二硫键与 J 链连接形成五聚体。IgG、IgD 和 IgE 常为单体,无 J 链。
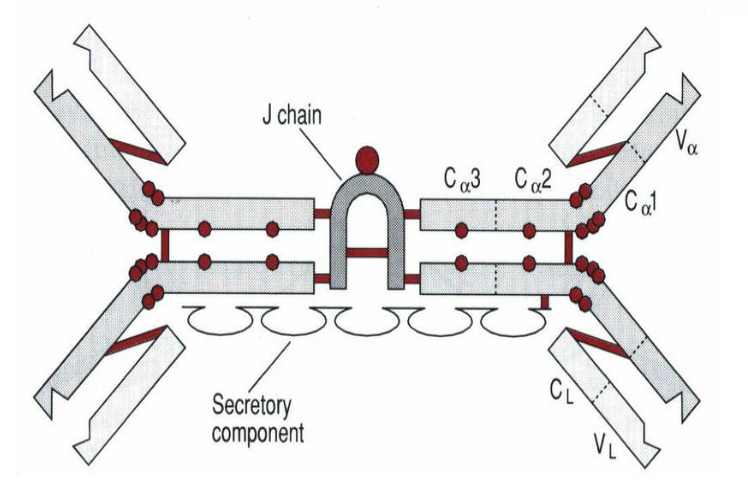
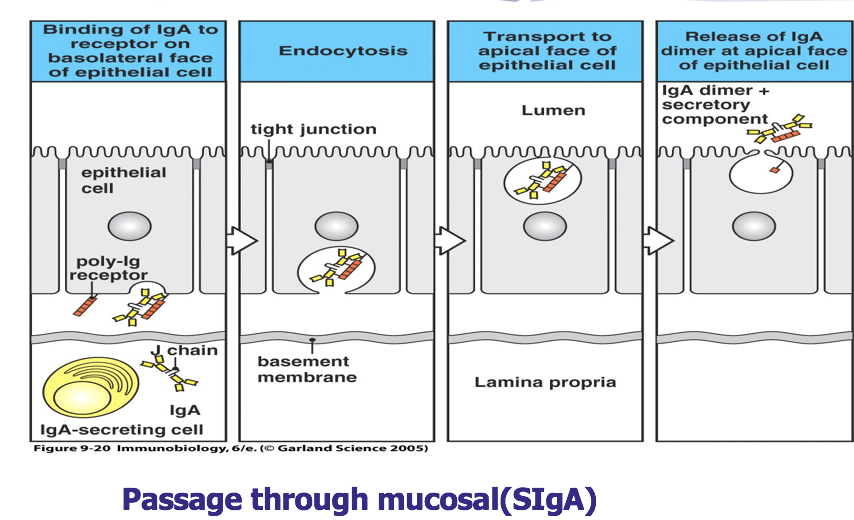
分泌型 IgA(SIgA)二聚体和 IgM 五聚体均由 J 链将其单体 Ab 分子连接为二聚体或五 聚体。分泌片(SP,图中橙色球组成的肽链)为一含糖肽链,是多聚免疫球蛋白受体(pIgR) 的胞外段,其作用是辅助 SIgA 由黏膜因有层,经黏膜上皮细胞转运,分泌到黏膜表面,并 保护 SIgA 链链区免遭蛋白水解酶降解。
将momomer结合到聚合物上

2.8.2 Secretory piece (分泌片)
分泌片(secretory piece,SP)又称分泌成分(secretory component,SC),是分泌 型 IgA 分子上的辅助成分,分子量约为 75kD,为含糖的肽链,由黏膜上皮细胞 合成和分泌,并以非共价键的方式结合于 IgA 二聚体上,与 J 链共同组成分泌型 IgA(SIgA)。分泌片具有保护 SIgA 的铰链区免受蛋白水解酶降解的作用,并介 导 SIgA 二聚体从黏膜下通过黏膜上皮细胞转运到黏膜表面。其功能是抗消化, 可保护 sIgA 不被分解。
- 分泌成分(Secretory component ,SC)是单个糖肽,MW为70KD。
- 由粘膜上皮细胞产生
- 功能:保护sIgA,抵抗额外分泌液中的蛋白水解。
2.10 水解片段
在一定调节下,可通过蛋白酶将Ig笑话成不同的片段,从而研究Ig的结构和功能,最常用的蛋白酶是木瓜蛋白酶和胃蛋白酶。
(一)木瓜蛋白酶水解片段
用木瓜蛋白酶可作用于 IgG 单体的铰链区二硫键,使其近 N 端切断,而得 到三个片段,其中两个片段完全相同,称为抗原结合片段(fragment of antigen binding,Fab),该段包含量了轻链和重链的可变区,能特异性地结合抗原;另一 片段可形成结晶,称为可结晶片段(fragment crystallizable,Fc),具有多种生物活 性,如激活补体、结合细胞、通过胎盘和黏膜等。Fab 由 VL、CL 和 VH、CH1 结构域组成,只与单个抗原表位结合(单价)。Fc 由一对 CH2 和 CH3 结构域组 成,含重链大部分 C 区,因而具有重链相应功能区的功能,是抗体与细胞表面 Fc 受体相互作用的部位,无抗原结合活性。
(二)胃蛋白酶水解片段
IgG 单体在胃蛋白酶作用下可在其铰链区二硫键近 C 端被切断,从而得到一 个具有双价抗体活性的抗原结合片段 F(ab’)2 和一些不具有任何生物活性的小分 子多肽碎片 pFc’。F(ab’)2 片段由 2 个 Fab 及铰链区组成,因此为双价,能够同 时结合两个抗原表位。由于 F(ab’)2 片段保留了相应抗原的生物学活性,同时又 避免了 Fc 段抗原性可能引起的副作用和超敏反应,因而被广泛用作生物制品。 从动物体内提取的抗毒素经胃蛋白酶水解后,可除去大部分 Fc 段,降低其免疫 原性,给人注射时可减少超敏反应的发生。pFc’片段由于最终被降解,因此不发 挥生物学作用。
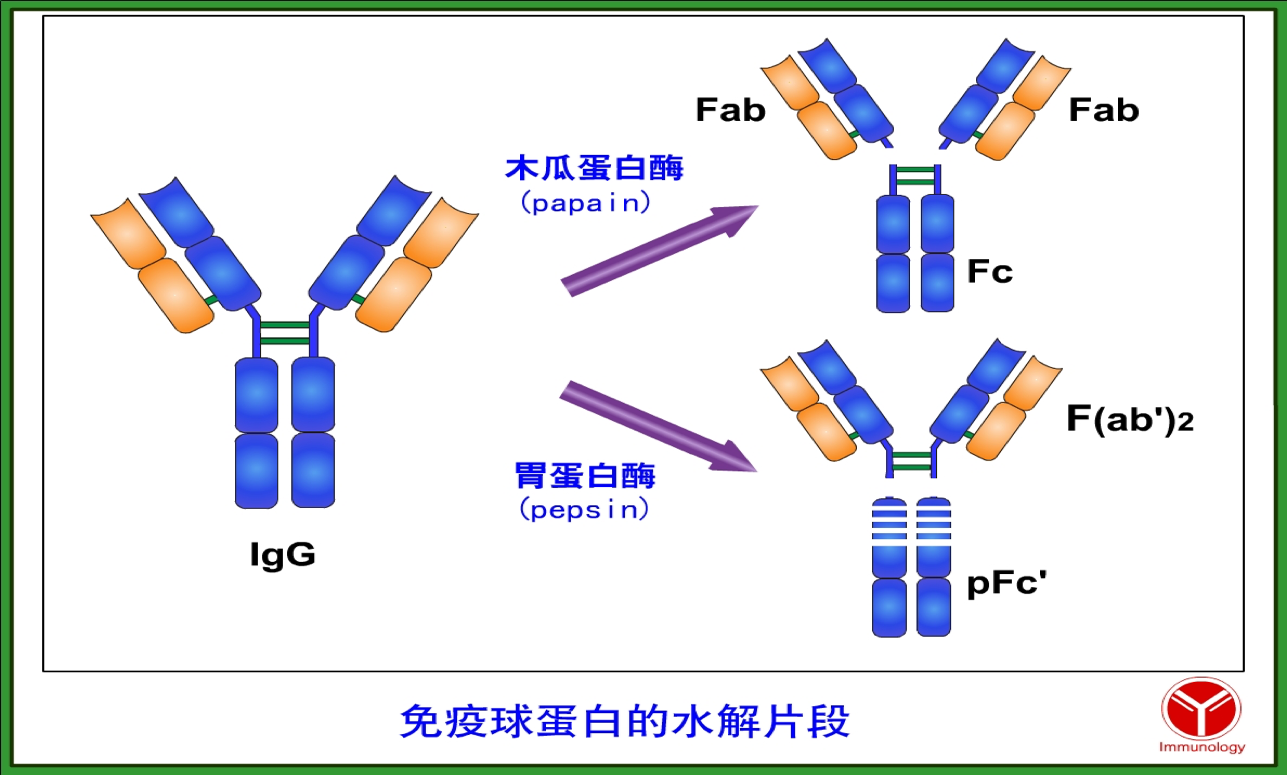
木瓜蛋白酶作用于铰链区二硫键所连接的两条重链近 N 端,将 Ab 裂解为 2 个完全相同的 Fab 段和一个 Fc 段。胃蛋白酶作用于铰链区二硫键所连接的两条重链近 C 端,将 Ab 水解 为 1 个大片段 F(ab’)2 和多个小片段 pFc’。
- Fab(fragment of antigen binding)
- Fc (fragment crystalizable)
比如白喉抗毒素, 破伤风抗毒素 胃蛋白酶消化后提纯,去掉FC 段而降低发生超敏反应
三、 抗体的多样性和免疫原性 Heterogeneity of immunoglobulins
由于不同抗原刺激 B 细胞所产生的抗体在特异性以及类型等方面均不尽相 同,因此抗体呈现出明显的多样性。自然界中抗原种类繁多,每种抗原分子结构 复杂,常含有多种不同的抗原表位。这些抗原刺激机体产生的抗体总数是巨大的, 包括针对各抗原表位的特异性的抗体,以及针对同一抗原表位的不同类型的抗体。 抗体的多样性是由免疫球蛋白基因重排决定并经抗原选择表现出来的,反映了机 体对抗原精细结构的识别和应答。
抗体既可与相应的抗原发生特异性结合,又因其本身具有免疫原性因而可激 发机体产生特异性免疫应答。其结构和功能的基础在于抗体分子中包含抗原表位。 这些抗原表位呈现三种不同的血清型:同种型、同种异型和独特型。
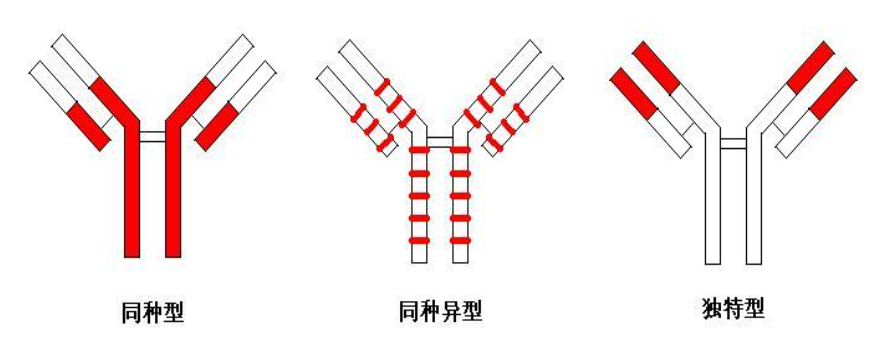
抗体分子存在 3 种不同的血清型:
- 同种型,指同一种属所有个体 Ab 分子共 有的抗原特异性标志,其表位存在于 Ab 的 C 区;
- 同种异型,指同一种属不同个 体间 Ab 分子所具有的不同抗原特异性标志,其表位广泛存在于 Ab 的 C 区;
- 独特型,指每个 Ab 分子所特有的、存在于 V 区的抗原特异性标志。
图中红色区域 代表抗体分子中三种血清型抗原表位所在部位。
Ig结同种型(Isotype )、异型( Allotype )、Idiotype
抗体作为Y形单元的一个或多个拷贝存在,由四条多肽链组成。 每个Y含有两个相同的重链拷贝和两个相同的轻链拷贝,它们的序列和长度不同。 Y形的顶部含有可变区,其与抗原上的表位紧密结合并特异性结合。
3.1 Isotype 同种型 (语言表述很不清楚,建议直接看图。。)
来源于不同种属的抗体分子对异种动物来说具有免疫原性,可刺激异种动物 (或人)产生针对该抗体的免疫应答。这种存在于同种抗体分子中的抗原表位即 为同种型。同种型因不同种属间重链和轻链的恒定区有较多的氨基酸不相同而具 有种属特异性但无个体特异性,例如,人与其他生物的免疫球蛋白同种型不同, 但不同人之间的免疫球蛋白同种型相同。因此,是同一种属所有个体抗体分子共 有拥有的抗原特异性标志,为种属型标志。
同种型(Isotypes )是一类Ig H的抗原特征(例如,链μ与γ不同)。一个物种的同种型可以诱导另一个物种的免疫反应。
在哺乳动物中,基于Y单元的数量和重链的类型,抗体被分为五种同种型:IgG,IgM,IgA,IgD和IgE。
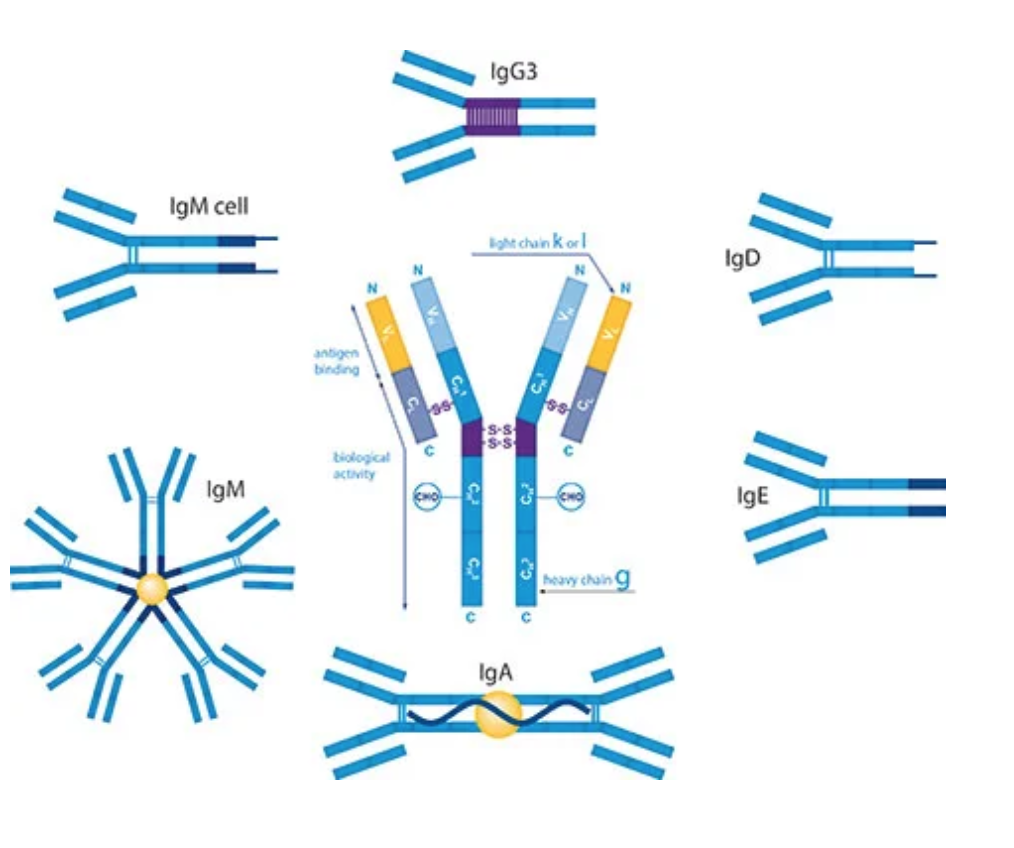
同种型的生物学特性,功能位置和处理不同抗原的能力不同:
| Isotype | Heavy chain | Light chain | MW (kDa) | Structure | Function |
|---|---|---|---|---|---|
| IgA1 IgA2 |
α1 α2 |
λ or κ | 150–600 | 单体 - 四聚体 | 大多数产生Ig。 存在于粘膜区域,例如肠道,呼吸道和泌尿生殖道,并且防止它们被病原体定植。 耐消化,分泌在牛奶中。 |
| IgD | δ | λ or κ | 150 | 单体 | 功能不清楚。 与B细胞发育中的IgM一起使用; 主要是B细胞结合 |
| IgE | ε | λ or κ | 190 | 单体 | 与过敏原结合并触发肥大细胞释放组胺,并参与过敏。 还可以防止寄生虫。 |
| IgG1 IgG2a IgG2b IgG3 IgG4 | γ1, γ2, γ3, γ4 | λ or κ | 150 | 单体 | 血清中的主要Ig。 提供基于对入侵病原体的免疫力的多数抗体。 中等补充修复剂。 IgG3可以穿过胎盘。 |
| IgM | μ | λ or κ | 900 | 五聚体 | 第一反应抗体。 表达在B细胞表面和具有非常高亲合力的分泌形式。 在有足够的IgG之前消除B细胞介导的免疫早期的病原体。 |
3.2 Allotype 同种异型
同一种属不同个体来源的抗体分子也具有各自的免疫原性,也可刺激不同个 体产生特异性免疫应答。这种存在于同种不同个体抗体中的抗原表位,称为同种 异型,是同一种属内不同个体间抗体分子因重链和轻链的恒定区有一至数个氨不 相同而具有的不同抗原特异性标志,为个体型标志。在 α 链、γ 链、κ 链发现有 数种至十多种遗传标志。
这是指物种内个体之间的遗传变异。
CH和CL上的一个或数个氨基酸残基的差异—遗传标志
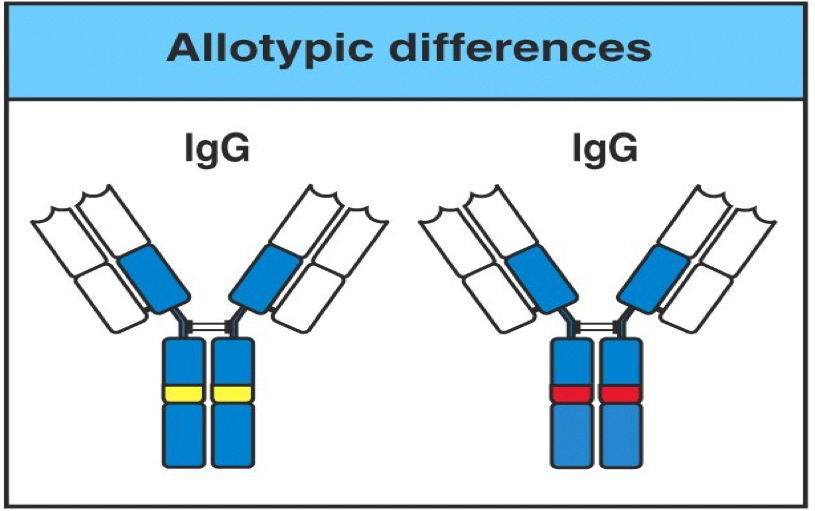
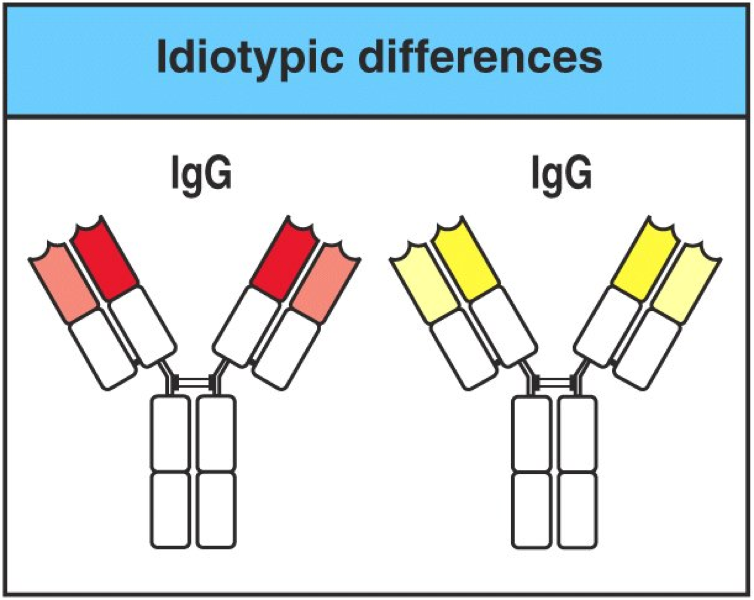
3.3 独特型 Idiotype
即使是同一种属、同一个体因不同抗原刺激而产生的抗体分子,其免疫原性 也不尽相同,称为独特型。独特型是每个抗体分子所特有的抗原特异性,其表位 被称为独特位(idiotope)。抗体分子可变区的氨基酸顺序和空间构型不同,每一 Fab 段约有 5~6 个独特位。独特型在异种、同种异体甚至同一个体内均可刺激产 生相应抗体,即抗特独特型抗体(anti-idiotype antibody,Aid 或 Ab2)。
V结构域的变异,特别是CDR中的变异,产生独特型(idiotype)。
四、抗体的功能
在体液免疫中,免疫球蛋白是发挥免疫效应的最重要的免疫分子。抗体的功 能与其结构密切相关。抗体分子的 V 区和 C 区的氨基酸组成及顺序的不同,决 定了它们功能上的差异;许多不同的抗体分子在 V 区和 C 区结构变化的规律性, 又使得抗体的 V 区和 C 区在功能上有各自的共性。V 区和 C 区的作用,构成了 抗体的生物学功能。
抗体可变区(V 区)和恒定区(C 区)的功能各异:
- V 区主要功能是特异性 结合抗原,从而阻断病原体入侵,发挥中和作用;
- C 区则在 V 区与特异性抗原 结合后,通过激活补体及与靶细胞表面 Fc 受体结合后,发挥调理作用、产生 ADCC 效应、介导超超敏反应和穿越胎盘等。
4.1 从功能上区分
4.1.1 特异性结合抗原
抗体最重要的功能是与相应抗原特异性结合,从而在体内介导多种生理和病 理效应。执行该功能的结构是抗体的 V 区,其中 CDR 在识别和结合特异性抗原 中起决定性作用。
抗体分子结构不同,因此结合抗原表位的数目也不相同。Ab 结合抗原表位 的个数称为抗原结合价。单体 Ab 可结合 2 个抗原表位,为双价;分泌型 IgA 为 4 价;五聚体 IgM 理论上为 10 价,但由于立体构型的空间位阻,一般只能结合 5 个抗原表位,故为 5 价。
B 细胞膜表面的 IgM 和 IgD 等 Ig 构成 B 细胞的抗原识别受体,能特异性识 别抗原分子。抗体的 V 区可以和抗原的表位特异性结合,从而封闭抗原的作用 位点,在体内发挥中和毒素、阻断病原入侵等免疫防御功能,还可以引起 C 区 变构,从而产生其他的生物学活性。在体外可发生各种抗原抗体结合反应,有利 于抗原或抗体的检测和功能的判断。
4.1.2 激活补体
抗体与相应抗原特异性结合后,可使抗体发生变构,而使其 CH2 和 CH3 结 构域内的补体结合位点暴露,补体成分与之结合,从而通过经典途径激活补体系 统,产生溶菌及溶细胞等多种补体的效应功能。其中 IgM、IgG1 和 IgG3 激活补 体的能力较强,IgG2 较弱。IgA、IgE 和 IgG4 本身难于激活补体,但形成聚合 物后可通过旁路途径激活补一系统。
4.1.3 结合 Fc 受体
抗体可通过其 Fc 段与细胞膜表面具有相应 Fc 受体(FcR)的细胞结合,产 生不同的生物学作用。IgG、IgA 和 IgE 的 Fc 受体分别称为 FcγR、FcαR 和 FcεR。
1.调理作用(opsonization) 细菌特异性的 IgG 抗体(特别是 IgG1 和 IgG3) 以其 Fab 段与相应细菌的抗原表位结合,以其 Fc 段与巨噬细菌或中性粒细胞表 面的 FcγR 结合,通过 IgG 的“桥联”作用,促进吞噬细胞对细菌的吞噬。

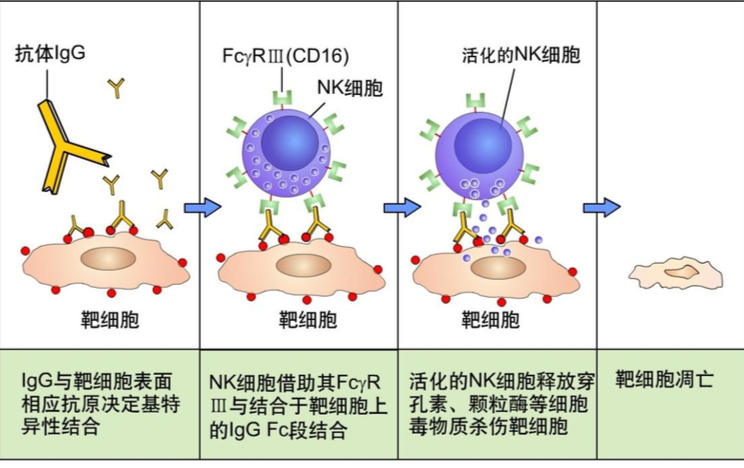
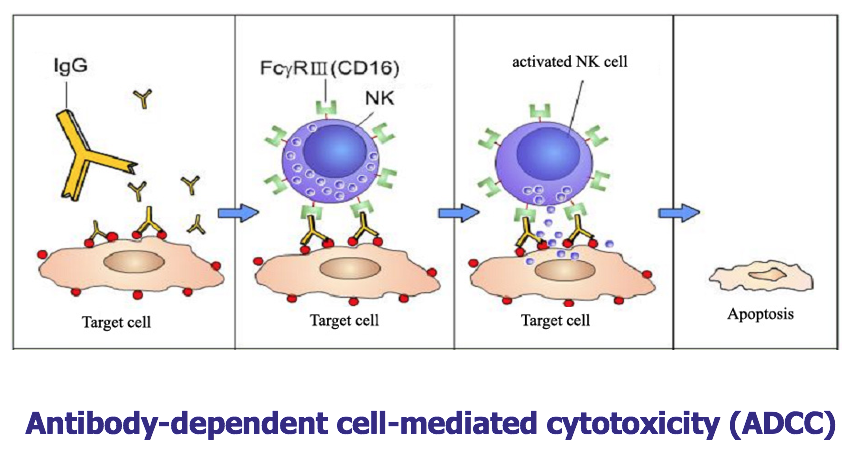
2.抗体依赖的细胞介导的细胞毒作用(antibody-dependent cell-mediated cytotoxicity,ADCC) IgG 抗体的 Fab 段结合病毒感染的细胞或肿瘤细胞表面的 抗原表位,其 Fc 段与自然杀伤细胞表面的 FcR 结合,自然杀伤细胞释放出穿孔 素、颗粒酶直接杀伤靶细胞。抗体与靶细胞上的抗原结合是特异性的,而表达 FcR 细胞的杀伤作用是非特异性的。
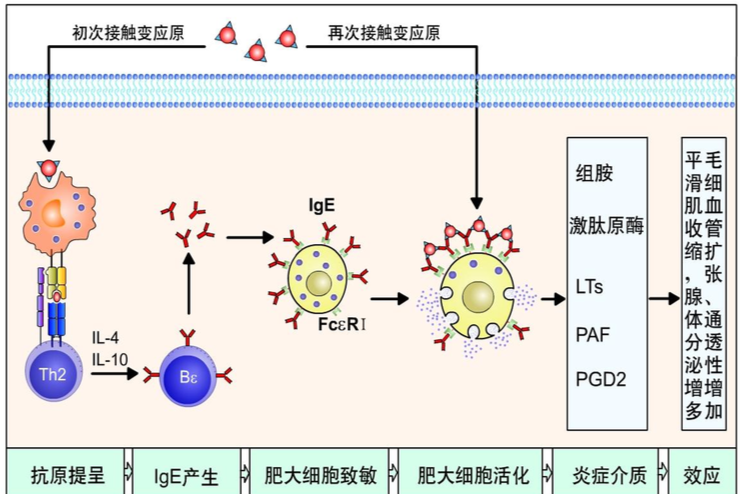
3.介导 I 型超敏反应 IgE 为亲细胞抗体,其 Fc 段可与肥大细胞和嗜碱性 粒细胞表面的高亲和力 IgEFc 受体(FcεRI)结合,并使其致敏。若相同变应原 再次进入机体与已致敏靶细胞表面特异性 IgE 结合,即可促使这些细胞合成和释 放生物活性物质,引起 I 型超敏反应。
在人类,IgG 是唯一能通过胎盘的免疫球蛋白。母体的 IgG 可经胎盘母体一 侧的滋养层细胞表达并输送蛋白,称为新生 Fc 段受体(neonatal FcR,FcRn)。IgG 可选择性与 FcRn 结合,从而转移到滋养层细胞内,并主动进入胎儿血液循环中。 IgG 穿过胎盘的作用是一种重要的自然被动免疫机制,对于新生儿抗感染具有重要意义。另外,分泌型 IgA 可被转运到呼吸道和消化道黏膜表面,是黏膜局部免 疫的最主要因素。
4.2 从区域来看功能
4.2.1 Functions of V regions
识别和结合抗原:
- HVR(CDR)
- 中和毒素(例如白喉,破伤风);
- 固定微生物;
- 中和病毒活性
Antibody:
- monomer(IgG,IgE)—2价
- dimer(sIgA) — 4价
- Petamer(IgM) — 5价
Neutralization 中和


4.2.2 Function of C regions (Fc portion)
- 激活补体( complement): IgM, IgG
- Ab( IgM、IgG )+Ag— C1q—-补体经典途径
- IgG4、IgA和IgE的凝聚物—-补体旁路途径
- Binding to Fc receptor of cells
- 调理作用(Opsonization): enhancement of Ag uptake by macrophages (MΦ)
- 抗体依赖性细胞介导的细胞毒性 (ADCC): NK cells.
- 参与I型超敏反应: IgE
- 通过胎盘(IgG, CH2) 和粘膜 (sIgA)

五、各类抗体的特性与功能
五类抗体中,IgG 和 IgM 以高浓度遍布全身,是全身性体液免疫反应的主要 效应分子。
5.1 IgG
婴儿出生 3 个月后开始合成,5 岁时达到成人水平。主要由脾和淋巴结中的 浆细胞合成和分泌,以单体的形式广泛存在于细胞外液中,是体液中含量最多的 一类抗体约占血清总 Ig 的确 75%~85%(表 4-1),在全身抗感染免疫中发挥最重 要的作用。人 IgG 有 4 个亚类,分别为 IgG1、IgG2、IgG3、IgG4。各亚类所占 比例:IgG1 为 60%~70%;IgG2 为 15%~20%;IgG3 为 5%~10%;IgG4 为 1%~7%。 IgG 主要由脾和淋巴结中的浆细胞合成,在血清中分解缓慢,半衰期约 20~23 天, 是再次免疫应答产生的主要抗体,其亲和力高,约有 50%的 IgG 存在于血清中, 其余 IgG 则广泛分布于各组织的细胞外液中,是机体抗感染的“主力军”。
IgG 有多能免疫球蛋白之称。IgG1、IgG3、IgG4 可穿过胎盘屏障,在新生 儿抗感染免疫中起重要作用。IgG1、IgG2、IgG3 的 CH2 能通过经典途径活化补 体,并可与巨噬细胞、NK 细胞表面 Fc 受体结合,发挥调理作用、ADCC 作用 等。人 IgG1、IgG2、IgG4 可通过其 Fc 段与葡萄球蛋白 A(SPA)结合,借此可 纯化抗体,并用于免疫诊断。某些自身抗体如抗甲状腺球蛋白抗体、抗核抗体, 以及引起 II、III 型超敏反应的抗体也属于 IgG。
特点:
- 单体分子;
- 四个亚类;
- 血清中含量最高(75%Ig);
- 半衰期最长(20~23天);
- 3~5岁达成人水平(8.0~17mg/ml);
- 可与SPA结合。
- 唯一能通过胎盘的Ig
功能:
- 修复补体-IgG1和IgG3修复补体; IgG4不能修复补体。
- 调理作用(Opsonization :巨噬细胞对Ag吸收的增强(MΦ)
- 抗体依赖性细胞介导的细胞毒性(ADCC):NK细胞
- 交叉胎盘IgG是唯一可以穿过胎盘的Ig类,它赋予新生儿高度的被动免疫力
5.2 IgM
主要由脾合成,分泌型 IgM 为五聚体由 J 链连接而成,分子质量最大,沉 降系数为 19S,因而被称为巨球蛋白。一般不通过血管壁,主要分布于血液中, 因此在防止菌血症中发挥着重要作用。IgM 占血清免疫球蛋白总量的 5%~10%, 血清浓度约为 1mg/ml。单体 IgM 以膜结合型(mIgM)表达于 B 细胞表面,构成 B 细胞抗原受体(BCR),只表达 mIgM 是未成熟 B 细胞的标志。
五聚体 IgM 含量 10 个 Fab 段,具有很强的抗原结合能力;含 5 个 Fc 段, 比 IgG 更易激活补体。IgM 也是个体出生后发生感染时产生最早的抗体,且半衰 期只有 5 天左右,故检测到血液中特异性 IgM 含量升高,可作为机体近期感染 的指标。在胚胎发育晚期的胎儿即能产生 IgM,脐带血中某些病毒特异性 IgM 水平升高提示胎儿有宫内感染(如风疹病毒或巨细胞病毒等感染)。IgM 也是初 次体液免疫应答中最早出现的抗体,是机体感染的“先锋队”;血清中检出病原 体特异性 IgM,提示新近发生感染,可用于感染的早期诊断。若机体缺乏 IgM, 当病原体侵入血流,易导致败血症的发生。IgM 不能通过胎盘,初乳中有 IgM, 可被婴儿吸收,出生后血清 IgM 很快增多,一岁时可达到成人量的 90%。ABO 天然血型抗体为 IgM 型;镶嵌于 B 细胞表面的 SmIgM 是 B 细胞的抗原受体。
- Penatmer 900kd巨球蛋白
- IgM是生命中合成的第一个Ig
- IgM是初次免疫应答中产生的第一个Ig。
- IgM是固定Ig的良好补体
- IgM也是一种良好的凝集Ig
- IgM是mIgM(IgD)的B细胞:
- 半衰期:5天

5.3 IgA
分为血清型和分泌型两种类型。血清型 IgA 为单体,主要存在于血清中,占 血清免疫球蛋白总量的 10%~15%。分泌型 IgA(SIgA)为二聚体,由 J 链连接, 含 SP,经黏膜上皮细胞分泌至外分泌液中,主要分布于呼吸道、消化道、泌尿 生殖道表面,以及唾液、泪液、乳汗和黏膜相关的分泌液中。SIgA 是外分泌液 中的主要抗体类别,参与黏膜局部免疫,通过与相应病原微生物结合,阻止病原 体黏附到细胞表面,从而在局部抗感染中发挥重要作用,是机体抗感染的“边防 军”。SIgA 在黏膜表面也有中和毒素的作用。新生儿易患呼吸道、胃肠道感染可 能与 IgA 合成不足有关。婴儿可从母亲初乳中获得 SIgA,是重要的自然被动免 疫。
- IgA:单体和分泌体(secretary)
- 血清IgA(IgA1)单体
- 分泌型IgA(IgA2)二聚体
- sIgA存在于泪液,唾液,初乳,粘液等分泌物中,对局部(粘膜)免疫有重要作用。
- IgA可以从母亲传播到新生儿
5.4 IgD
正常人血清中 IgD 含量最低(约 30μg/ml),仅占血清免疫球蛋白总量的 0.3%。 可在个体发育的任何时间产生。五类 Ig 中,IgD 的铰链区较长,易被蛋白酶水 解,故其半衰期很短(仅 3 天)。IgD 分为血清型 IgD 和膜结合型 IgD。前者的 生物学功能仍不清楚;后者是 B 细胞分化发育成熟的标志,未成熟 B 细胞仅表 达 mIgD,成熟 B 细胞可同时表达 mIgM 和 mIgD,称为初始 B 细胞(naive B cell); B 细胞活化后其表面的 mIgD 逐渐消失。
B 细胞表面的 IgD(SmIgD)可作为 B 细胞成熟的重要标志,同时也是 B 细 胞的抗原受体。
- 血清中IgD水平较低; 它在血清中的作用仍不清楚。
- IgD主要存在于naive B细胞表面,其中它作为抗原的受体起作用。 3.膜IgD在活化或记忆B细胞中消失
5.5 IgE
分子量为 160kD,是种系进化中最晚出现的免疫球蛋白。以单体形式存在于 血清中。是正常人血清中含量最少的一类 Ig,血清浓度极低,约为 3×10-4mg/ml。 主要由黏膜下淋巴组织中的浆细胞分泌。IgE 的重要特征在于它是一类亲细胞抗 体,其 CH2 和 CH3 结构域可与肥大细胞、嗜碱性粒细胞上的高亲和力 FcεR1 结 合,当结合再次进入机体的抗原可引起 I 型超敏反应。此外,IgE 可能与机体抗 寄生虫免疫有关。在过敏性疾病和寄生虫感染时,患者血清中 IgE 含量可显著增 高。IgE 为亲细胞抗体,其 Fc 与肥大细胞、嗜碱性粒细胞结合,参与 I 型超敏反 应。
- IgE是最不常见的血清Ig。 它甚至在与抗原相互作用之前与嗜碱性粒细胞和肥大细胞上的Fc受体非常紧密地结合。
- 参与过敏反应。
- IgE还有助于通过嗜酸性粒细胞清除寄生虫(例如蠕虫)。

六、抗体种类
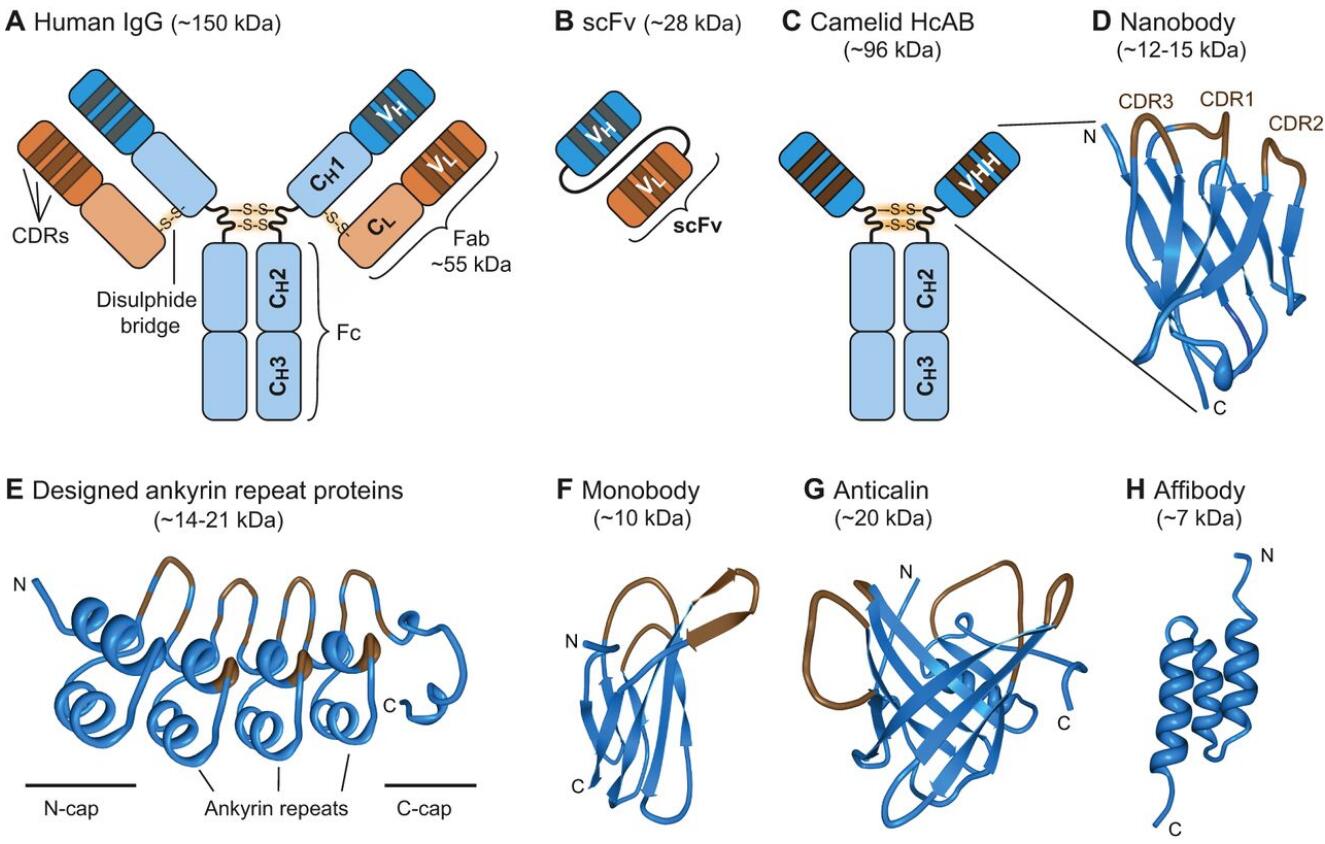
The structure and size of selected protein binder scaffolds. The structures of different protein binder scaffolds are shown; variable regions are marked in brown. (A) Conventional human immunoglobulin (IgG) antibody, consisting of two heavy (blue) and two light (orange) chains connected by disulphide bridges (S-S). Both chains consist of several constant domains (CL and CH1-CH3) and the variable domains (VL and VH). The variable domains contain the complementarity-determining regions (CDRs), which are the three hypervariable domains that determine antibody-binding specificity. (B) The single-chain variable fragment (scFv) is a synthetic fusion of the VH and VL domains via a linker. (C) Camelid heavy-chain antibodies (HcABs) are made up of only two heavy chains, each of which consists of two constant (CH2 and CH3) domains and a single variable (VHH) domain. (D) Structure of an anti-GFP nanobody (vhhGFP4, PDB ID:3OGO; Kubala et al., 2010). The positions of the three CDRs are highlighted. (E) Structure of the AR_3a DARPin (PDB ID:2BKK; Kohl et al., 2005). DARPins typically consist of constant N- and C-capping repeats motives (N-/C-cap), encapsulating two or three variable ankyrin repeats (three are shown here). (F) Structure of the monobody ySMB-1 (PDB ID: 3QHT; Gilbreth et al., 2011). (G) Anticalin US7 structure (PDB ID: 4MVI; Rauth et al., 2016). (H) Structure of the ZHER2 affibody (PDB ID: 2KZJ; Eigenbrot et al., 2010).
七、基因结构
Ig基因及其重排
Ig的V区位于重链(H链)和轻链(L链)的N端。编码H链V区的基因包含V、D、J三组基因片段;编码L链V区的基因包含V、J两组基因片段。
- H链的V基因编码H链V区的CDR1及CDR2互补决定区、D基因编码大部分CDR3互补决定区、J基因编码剩余部分CDR3以及V区和C区的连接结构;
- L链的V基因编码H链V区的CDR1、CDR2和大部分CDR3互补决定区、J基因编码剩余部分CDR3以及V区和C区的连接结构。 由以上信息可以看出,Ig V区是由V、(D)、J基因片段共同翻译出来的,V、(D)、J基因片段重排共同改变了Ig可变区的结构,从而影响BCR的多样性。
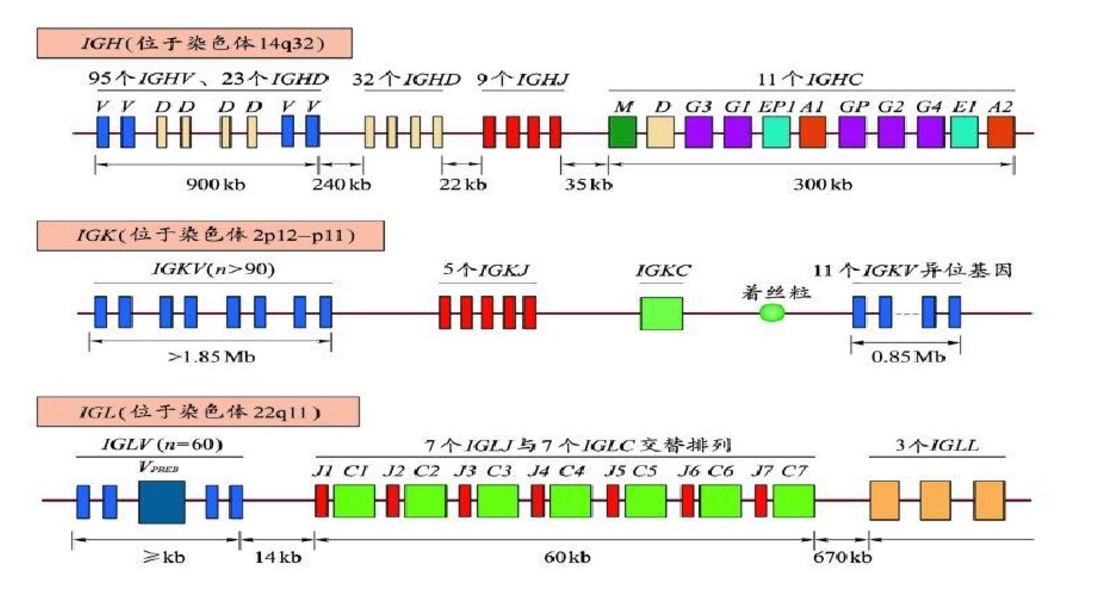
- Ig基因群:V , D ,J ,C
- IGH : IGHV, IGHD, IGHJ, IGHC
- IGK/IGL: IGKV/IGLV, IGKJ/IGLJ, IGKC/IGLC
八、抗体的产生
抗体在疾病的诊断、免疫防及其基础研究中被广泛应用,人们对抗体的需求 也随之增大。人工制备抗体是大量获得抗体的有效途径。以特异性抗原免疫动物, 制备相应的抗血清,是早年人工制备多克隆抗体的主要方法。1975 年,KÖhler 和 Milstein 建立的单克隆抗体(单抗)技术,使得规模化制备高特异性、均一性 抗体成为可能。但鼠源性单抗在人体反复使用后出现的人抗鼠抗体(HAMA)反 应,很大程度上限制了单抗的临床应用。近年,随着分子生物学的发展,人们已 可通过抗体工程技术制备基因工程抗体,包括人-鼠嵌合抗体、人源化抗体或人 抗体等。
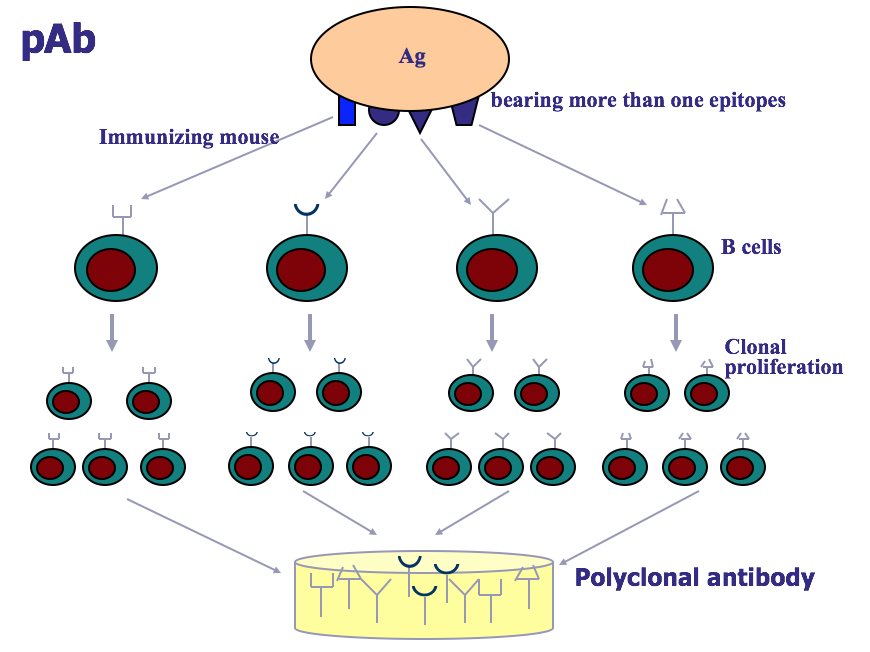
8.1. 多克隆(Polyclonal )抗体
天然抗原常含多种抗原表位。以该抗原刺激机体免疫系统,可使多种具有相 应抗原受体的 B 细胞活化增殖发生免疫应答,分泌多种针对不同抗原表位的抗 体。由不同 B 细胞产生的、针对多种抗原表位的混合抗体称为多克隆抗体 (polyclonal antibody,pAb)。获得多克隆抗体的途径主要有动物免血清、恢复期 中层得血清或免疫接种人群。其优点是:作用全面,具有中和抗原、免疫调理、 介导补体依赖的细胞毒作用(CDC)、ADCC 等重要作用,来源广泛、制备容易; 缺点是:特异性不高、易发生交叉反应,不易大量制备,从而应用受限。
8.2. 单克隆( Monoclonal )抗体
KÖhler 和 Milstein 将可产生特异性抗体但短寿的 B 细胞与无抗原特异性但长 寿的骨髓瘤细胞融合,建立了可产生单克隆抗体的 B 淋巴杂交瘤细胞和单克隆 抗体技术。通过该技术融合形成的杂交细胞系即杂交瘤,既有骨髓瘤细胞大量扩 增和永生的特性,又具有免疫 B 细胞合成和分泌特异性抗体的能力。每个杂交 瘤细胞由一个 B 细胞与一个骨髓瘤细胞融合而成,而每个 B 细胞克隆仅识别一 种抗表位,故经筛选和克隆化的杂交瘤细胞仅能合成及分泌抗单一抗原表位的特 异抗体。这种由单一杂交瘤细胞产生,针对单一抗原表位的特异性抗议体,称为 单克隆抗体(monoclonal antibody,mAb),具有结构均一、纯度高、特异性强、 效价高、易体外大量制备和强化等特点,已广泛应用于医学及生物学各领域,如 细胞因子、肿瘤细胞表面抗原受体、激素等的检测。
Georeges Kohler and Cesar Milstein, 1984, monoclonal antibody

Human Monoclonals?
It would be very nice to have available human monoclonals - much more useful therapeutically
Problems:
- Hard to get antigen-primed B cells
- In vitro primed
- SCID mice
- Knock-out mice
- No good immortal partners
- Can sometimes transform antibody-secreting B cells (EBV)
8.3. 基因工程抗体
基因工程抗体是应用 DNA 重组和蛋白质工程技术,按照人们的意愿在基因水平 上对免疫球蛋白分子进行切割、拼接或修饰,重新组装成的新型抗体分子。基因 工程抗体保留了天然抗体的特异性和主要生物学活性,去除或减少了无关结构, 并赋予抗体分子新的生物学活性,因此该抗体具有更广泛的应和前景。通过基因 工程技术可制备的基因工程抗体有:人-鼠嵌合抗体、人源化抗体、双特异性抗 体、小分子抗体及人抗体等。
- Chimeric Ab
- Mouse V gene + human C gene vector expression
- CDR-grafted Ab
- Mouse CDR gene human framework region
- Bispecific antibody

参考资料
- https://wenku.baidu.com/view/a14650ef81c758f5f61f679d.html
- https://zhidao.baidu.com/question/1737599212496478627.html
- Protein binders and their applications in developmental biology
- https://www.abcam.com/protocols/antibody-structure-and-isotypes
- https://en.wikipedia.org/wiki/Immunoglobulin_light_chain
- 《Immunoglobulin And Antibody》,Xiaojian wang, Ph.D. Institute of Immunology ZJU School of Medicine
- 《医学免疫学》 新乡医学院 课件
