【1.7.1】免疫细胞膜分子--CD分子与粘附分子
一、人白细胞分化抗原
1.1 人白细胞分化抗原和 CD 的概念
人白细胞分化抗原(human leukocyte differentiation antigen,HLDA)主要是指造血干细 胞在分化成熟为不同谱系、各谱系分化的不同阶段以及细胞成熟后活化过程中所表达的细胞 表面分子。目前已知白细胞分化抗原不仅存在于白细胞表面,还广泛分布于红细胞、血小板、 血管内皮细胞、上皮细胞、成纤维细胞和神经内分泌细胞等多种细胞表面。HLDA 多为跨膜 糖蛋白,少数为碳水化合物;根据胞外区结构特点可将其分为
- 免疫球蛋白超家族、
- 肿瘤坏死 因子超家族、
- 细胞因子受体家族、
- C 型凝集素超家族和选择素家族。
Köhler 和 Milstein1975 年建立的单克隆抗体技术极大地推动了人们对 HLDA 的研究。 国际专门命名机构将来自不同实验室的单克隆抗体所识别鉴定的同一种分化抗原归为同一 个分化群(cluster of differentiation),简称 CD。单克隆抗体及其识别的相应抗原表位通常共 用一个 CD 编号,即一个 CD 编号既可代表某种单克隆抗体,又可代表该种单克隆抗体识别 鉴定的细胞膜表面分子。目前人白细胞分化抗原多数已有 CD 编号,有关 CD 分子编号 (CD1~CD363)及其主要特征参见本书附录II。
1.2 人白细胞分化抗原的分类和相关分子的主要功能
根据功能特征可将人白细胞分化抗原大致分为受体和黏附分子两类:
- 其中参与识别和信 号转导的受体分子主要包括 TCR-CD3 复合体、BCR-Igα/Igβ 复合体、TCR/BCR 辅助受体、 NK 细胞杀伤活化受体、模式识别受体如 Toll 样受体(TLRs)、IgGFc 受体、补体受体、细 胞因子受体和死亡受体;
- 可介导细胞间或细胞与细胞外基质间相互作用和信号转导相关的黏 附分子主要包括共刺激分子、归巢受体和地址素等。
细胞表面与其识别和黏附相关的分子及其主要作用



PRR(patternrecognitionreceptor,模式识别受体);CKR(cytokinesreceptors, 细胞因子受体 );TNFR1(Tumor necrosis factor 1, 肿瘤坏死因子 1);SelX(Sialy leveisX, 唾液酸化的路易斯寡糖 X); LFA-1(lymphocyte function associated antigen -1, 淋巴细胞功能相关抗原-1 );ICAM-1(intercellular adhesion molecules-1,细胞 间黏附分子-1); GlyCAM-1(glycosylation-dependentcelladhesionmolecule-1,糖基化依赖的细胞黏附分子 -1);PNAd(peripheral lymphnode vascular addression,外周淋巴结血管地址素)
二、黏附分子
细胞黏附分子(cell adhesion molecules,CAM)是介导细胞间或细胞与细胞外基质 (extracellular matrix ECM)间相互作用的分子,简称黏附分子。黏附分子为跨膜蛋白,以受 体配体结合形式发挥作用;参与细胞的识别活化、增殖分化、信号转导和迁徙移动。他们 在免疫应答、炎症反应、肿瘤转移和创伤愈合等病生理过程中发挥重要作用。黏附分子属白 细胞分化抗原,多数已有 CD 编号;根据黏附分子结构特点可将其分为免疫球蛋白超家族、 整合素家族、选择素家族和钙黏蛋白家族等。
2.1 免疫球蛋白超家族
免疫球蛋白超家族(immunoglobulin superfamily, IgSF)是一类结构和氨基酸组成与 Ig 可变区或恒定区结构域相类似的同源蛋白分子。IgSF 成员在免疫细胞众多膜分子中所占比 例最大,其种类繁多、分布广泛、功能各异;主要参与淋巴细胞对抗原的识别,免疫细胞间 的相互作用和细胞活化信号的转导。表 7-2 摘要介绍在 T 细胞与 APC 结合相互作用过程中 所涉及的 IgSF 黏附分子及其主要功能。
T 细胞与 APC 结合相互作用过程中涉及的黏附分子及其主要功能

ICOS(inducible costimulator, 诱导性共刺激分子);PD-1(programmed death-1, 程序性死亡-1);PDL- 1(programmed death ligand-1, 程序性死亡配体-1)
2.2 整合素家族
整合素家族(integrin family)成员是由 α 和 β 两条肽链组成的异二聚体分子,因其主要 介导细胞与细胞外基质黏附,使细胞得以附着形成整体(integration)而得名。整合素家族中 至少有 18 种链,8 种链,共有 24 种组合形式。根据链的不同,可将整合素家族分 8 组: 在同一组中,β 链相同而 α 链不同;通常大多数 α 链只结合一种 β 链,有些 α 链可结合多种 链,分组不绝对严格。整合素在体内分布广泛,一种整合素可分布于多种细胞;同一种细 胞也可表达多种整合素。整合素分子表达水平受细胞活化和分化状态的影响,某些整合素的 表达具有显著的细胞类型特异性。整合素家族1~2 组中某些成员的分子结构、分布、配体 和主要功能如表所示。

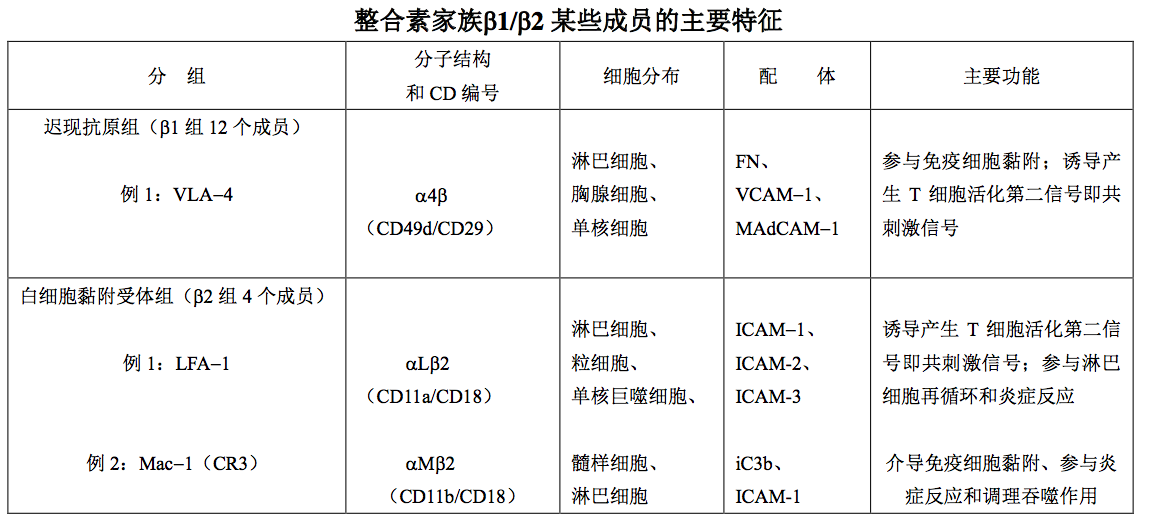
注:
VLA-4(very late antigen-4,迟现抗原-4);FN(fibronectin,纤连蛋白);VCAM1(vascular cell adhesion molecule-1,血管细胞黏附分子-1);MAdCAM-1(mucosal addressin cell adhesion molecul-1,黏膜地址素细 胞黏附分子-1);LFA-1(lymphocyte function associated antigen-1,淋巴细胞功能相关抗原-1);ICAM-1,2,3 (intercellular adhesion molecule-1,2,3,细胞间黏附分子-1,2,3)。
2.3 选择素家族
选择素家族(selectin family)成员为跨膜分子,其胞膜外区均由 C 型凝集素样(CL) 结构域、表皮生长因子(EGF)样结构域和补体调节蛋白(CCP)结构域组成:其中胞外区 CL 结构域是与相应配体结合的部位,其胞质区与细胞骨架相连。
选择素家族有三个成员,即白细胞选择素(leukocyte-selectin,L-选择素)、内皮细胞选 择素(endothelium-selectin,E-选择素)和血小板选择素(platelet-selectin,P-选择素)。选择 素分子识别的配体主要是表达于白细胞和内皮细胞表面的某些寡糖基团和糖蛋白,如唾液酸 化的路易斯寡糖(sialylLewisx,sLex)和糖基化依赖的细胞黏附分子(glycosylation-dependent cell adhesion molecule-1,GlyCAM-1)等。他们在淋巴细胞归巢、炎症以及白细胞与血管内皮细 胞黏附过程中发挥重要作用。三种选择素的分布、配体及其主要功能如表所示。



2.4 黏附分子的主要功能
黏附分子通常以受体-配体结合方式发挥作用,参与免疫应答、炎症反应和肿瘤转移等 一系列重要生理和病理过程,摘要简介如下。
1 介导 T 细胞与 APC 结合启动适应性免疫应答
抗原提呈细胞表面黏附分子与 T 细 胞表面相应黏附分子互补结合是启动适应性免疫应答的关键步骤,其过程简述如下:T 细胞 进入外周免疫器官后首先通过表面 LFA-1 和 LFA-2 等粘附分子与 APC 表面 ICAM-1 和 LFA- 3 等相应黏附分子松散结合,使二者发生滚动和可逆性粘附从而为 T 细胞表面 TCR-CD3 复 合体对 APC 表面相应抗原肽-MHC 分子复合物的特异性识别创造了条件。
T 细胞通过表面 TCR-CD3 复合体从 APC 表面众多抗原肽-MHCII/I类分子复合物 (pMHC)中挑选出相应抗原肽-MHCII/I类分子复合物,并与之特异性结合后诱导产生 T 细胞活化第一信号。上述活化信号可使 T 细胞和 APC 表面某些黏附分子构象改变聚集在 TCR-pMHC 周围、形成免疫突触(immunologycal synapse),并由此导致 T 细胞表面黏附分 子 CD28、LFA-2、LFA-1 与 APC 表面黏附分子 B7、LFA-3、ICAM-1 间的亲和力显著增强, 成为诱导产生 T 细胞活化第二信号的共刺激分子,从而有效激活抗原特异性 T 细胞启动适 应性免疫应答。
2 介导血管内中性粒细胞向感染炎症部位迁移
血管内中性粒细胞向感染炎症部位迁 移过程简述如下: 1)感染部位巨噬细胞被病原体激活后产生的 IL-1 和 TNF-α 等促炎细胞 因子可使相邻毛细血管内皮细胞活化,表达 E-选择素、整合素配体 ICAM-1 及膜型趋化因子 mIL-8 和分泌型 IL-8。2)中性粒细胞通过表面唾液酸化的路易斯寡糖(Sialy LewisX,Slex) 与血管内皮细胞表面 E-选择素松散结合发生滚动黏附,可使中性粒细胞表达 IL-8R,并通过 对内皮细胞表面膜型 IL-8 的结合起到制动作用:同时可刺激中性粒细胞,使其表面整合素 LFA-1 表达上调和活化;使血管内皮细胞表面 ICAM-1 表达上调。 3)中性粒细胞通过表面 Slex 、IL-8R、高亲和力 LFA-1 与内皮细胞表面相应 E-选择素、mIL-8、ICAM-1 结合形成稳 定黏附。4)上述变化导致中性粒细胞骨架重组,使其趴伏在内皮细胞表面形成紧密黏附, 产生以下两种作用:1刺激内皮细胞连接处一种称之为 VE-钙粘素复合物(VE-cadherin complex)的蛋白发生短暂而可逆性解离(disruption),从而导致内皮细胞间隙开放,为中 性粒细胞外渗提供了“方便之门”。2中性粒细胞表面 LFA-1 与内皮细胞表面 ICAM-1 之间亲 和力显著降低;与此同时,在感染部位巨噬细胞和血管内皮细胞分泌的高浓度 IL-8 等趋化 因子作用下,中性粒细胞从血管内皮细胞间隙渗出,进入感染部位发挥抗感染免疫作用。血 管内单核细胞和 T 淋巴细胞也能以类似于上述中性粒细胞外渗迁移方式进入感染部位发挥 作用。

中性粒细胞对血管内皮的黏附和渗出过程
3 介导淋巴细胞归巢
淋巴细胞归巢(lymphocyte homing)是指初始 T、B 淋巴细胞离 开中枢免疫器官后,经血液循环定向迁移到外周免疫器官和淋巴组织的过程。T 淋巴细胞表 面与其归巢相关的黏附分子又称淋巴细胞归巢受体(lymphocyte homing receptor,LHR), 主要包括 L-选择素和、LFA-1。淋巴细胞归巢受体识别结合的配体表达于内皮细胞表面称之 为血管地址素(vascular addression), 主要包括表达于淋巴结高内皮小静脉内皮细胞表 面的外周淋巴结血管地址素(peripheral lymph node vascular addression,PNAd,CD34)、 糖基化依赖的细胞黏附分子 1(glycosylation-dependent cell adhesion molecule 1, GlyCAM-1)、ICAM-1、2 和表达于派氏小结高内皮小静脉和粘膜固有层小静脉内皮细胞表面 的黏膜地址素细胞黏附分子-1(mucosal addreession cell adhesion molecule-1,Mad CAM-1)。研究证实,生理条件下淋巴结高内皮小静脉内皮细胞表面 CD34、GlyCAM-1、ICAM- 1、2等黏附分子和膜型次级淋巴组织趋化因子(Secondary lymphoid tissue chemokine, SLC, 即 CCL21 )表达与淋巴结深皮质区中树突状细胞对内皮细胞的激活作用密切相关,即 树突状细胞通过表面 LTα1β2 三聚体与高内皮小静脉内皮细胞相应受体(LTβR)结合,可 诱导高内皮小静脉内皮细胞成熟,使之表达上述黏附分子及膜型 SLC 和分泌型 SLC、即 CCL21。
初始 T 细胞与高内皮小静脉内皮细胞间的相互作用如图 7-4 所示:1首先通过其表面 L-选择素与血管内皮细胞表面 GlyCAM-1/CD34 松散结合,沿血管壁发生滚动黏附;2进而 通过表面趋化因子受体(CCR7)与内皮细胞表面膜型次级淋巴组织趋化因子 (mSLC)、即膜型 CCL21 结合,诱导 T 细胞表面 LFA-1 活化,使之与血管内皮细胞表面相应黏附分子 ICAM-1 结合,导致 T 细胞与血管内皮细胞紧密黏附;3在淋巴结深皮质区高内皮小静脉内 皮细胞和成纤维网状细胞(fibroblastic reticular cells,FRC)分泌的 CCL21 和 CCL19 作用下,初始 T 细胞穿过血管内皮细胞进入淋巴结深皮质区,参与适应性免疫应答。
初始 B 细胞也能通过与初始 T 细胞类似的作用方式进入淋巴结深皮质区;但初始 B 细 胞低表达 CCR7 而高表达 CXCR5,因此在滤泡树突状细胞(FDC)分泌的 B 淋巴细胞趋化因 子 1 (B lymphocyte chemokine 1,BLC-1)即 CXC13 作用下,能够从深皮质区继续迁移进 入淋巴滤泡,参与适应性体液免疫应答。

T 淋巴细胞归巢过程示意图
参考资料
- 《医学免疫学》课件 新乡医学院 宋向凤、张国俊、徐春阳、孙爱平、孙书明、赵铁锁等老师
