【2.6】ig-like domain
序列基序(motif)是广泛存在的核苷酸或氨基酸序列模式,具有或可用于推测生物学意义。
Ig-like motif具有如下的特点:
- 所有Ig-like结构域都参与结合功能
- 经典的Ig-like结构域由7到10个β链组成,分布在两个sheet之间
- Ig-like结构域的一般形状是很保守的,但由于环的高度可变性,它们的大小可能有很大差异
- 可以根据β链的数量对Ig样结构域进行分类: C1-type、C2-type、V-type、H-type
更多的信息的理解,见下面的参考资料中prosite的说明
先根据结构来定义ig-like; 然后基于这样的结构,来分析序列,看那些序列特征符合这种结构,符合这样结构特征的定义为一个Motif; 基于这样的motif,就可以预测序列是否为ig-like
文献内容
脊椎动物的免疫系统是非常了不起的,因为它几乎可以识别任何外来物质或抗原,但除了在自然免疫导致的极少数情况下,它们不会对自身产生反应。 在过去的20年中,已经充分表征了抗原被B细胞抗原受体(Ig)和T细胞抗原受体(TcR)识别的机制。 这两组蛋白质都是由Ig样结构域构建的。 variable or V结构域提供与外来抗原范围反应的特异性,并且constant or C结构域提供效应子功能。 一个引人入胜的问题是如此复杂的系统是如何演变的,这是Strong等人在本期PNAS论文中提出的一个主题。
Igs和TcRs的结构域结构如图1所示。V和C结构域都具有相似的结构,称为Ig折叠,它由反平行β-strand排列成两个由二硫键连接的片段形成(图2)。 V结构域可以与C结构域区分开来,因为它们显示出与抗原识别相关的可变性,并且结构域也更长,其中两个额外的链被容纳在结构域的中间(图2中的C’和C“)。由TcR(例如,αβ)和Igs(重链和轻链)中的两条链构成。另外的链与Ig和TcR相关,其介导向细胞质的信号传导.CD79a和CD79b与膜Ig结合并含有参与信号转导的免疫受体酪氨酸激活基序(immunoreceptor tyrosine activation motif, ITAM).TcR异二聚体与CD3链相关,包括含有ITAM基序的ζη。允许Ig和TcR结合这种多样化蛋白质的变异性主要取决于在域顶部的每个链中的三个环(图2中用红色着色),其显示出特定的可变性(高变区)。
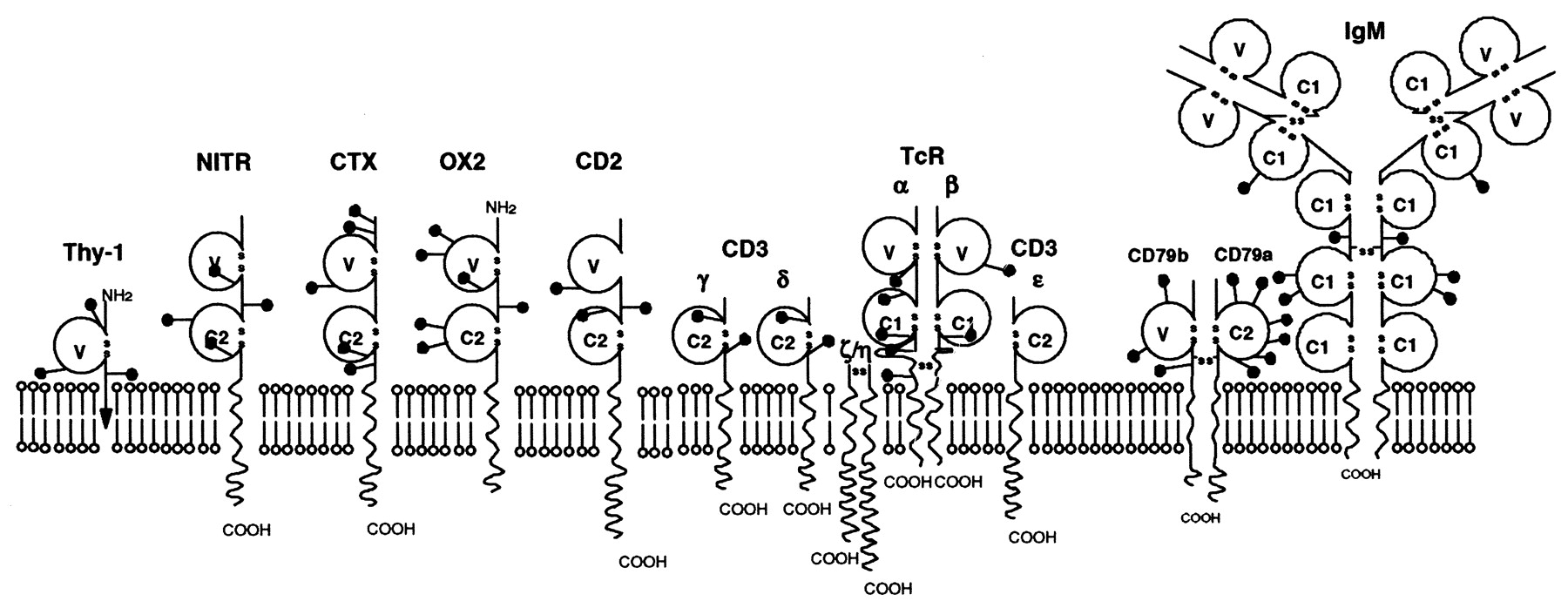
图1 Blob图说明含有IgSF结构域的各种蛋白质,包括抗原受体。 IgSF结构域用空心圆圈表示,并根据序列模式和大小标记为V,C1或C2(7)。 棒棒糖符号表示N-连接糖基化位点的大致位置。 Thy-1中的箭头表示存在糖基 - 磷脂酰肌醇锚。
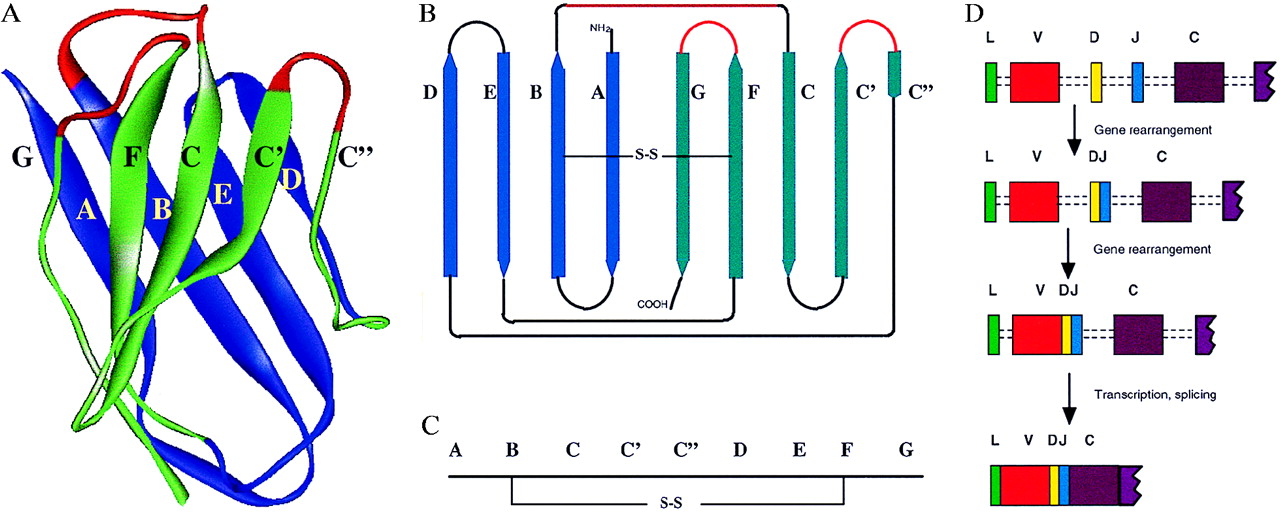
图2 Ig折叠。 (A)人骨髓瘤(Protein Database 7fab)的重链V区的X射线晶体学结构的带状图。 (B)形成两片折叠的股线的示意图。 在A和B中,ABED薄片为蓝色,GFCC’C“薄片为绿色。构成高变区的三个循环用红色表示。(C)B中链的线性表示。注意C域缺乏 额外的循环C’和C“。 C2结构域缺乏C“链。(D)产生单个TcRβ或重链的方案,其中选择特定的V区,D和J区段并重组以形成VDJ区段。转录后,将内含子拼接出来以得到。为简单起见,不包括编码C区和跨膜的其他外显子。
variability是如何产生的? V结构域通过体细胞中的重组从基因区段的多个拷贝构建。大部分结构域由V区段编码,并与D区段和J区段中的一个组合形成VDJ外显子,然后在转录后剪接至C结构域(图2D)。这种连接不是完全精确,也引入了所谓的N-区域多样性(不同抗原受体之间存在一些细节差异;参考文献2)。 J区构成最终β-链的大部分(图2中的G)并且具有特征序列模式(参见参考文献1的图3C)。这种奇妙复杂的抗原识别分子系统是如何在进化过程中产生的?当Igs在20世纪60年代中期进行测序时,很快就发现它们是由具有相似序列模式的区域构成的,这表明V和C结构域是通过基因复制进化的(3,4)。 MHC抗原和β2微球蛋白的序列数据表明Ig-like序列可能存在于与免疫应答相关的其他蛋白质中。当脑抗原Thy-1和随后的数百种其他蛋白质被发现含有Ig相关序列时,很明显在自适应免疫系统的重排受体之前存在Ig折叠(5-7)。这些序列区域通常被称为Ig超家族(Ig superfamily, IgSF)或Ig-like结构域,以将它们与Igs本身区域区分开来。结果表明,Ig折叠对于介导相互作用特别有益,并且被广泛用于例如神经系统中,而不是用于抗原识别的特殊事物。根据序列模式和总长度(7-9),可以将IgSF结构域分为V,C1,C2和I。应当注意,IgSF V结构域显示与Ig本身的V结构域的序列相似性,但它们没有显示通过重排产生的Ig或TcR V结构域中发现的变异性(参见上文)。因此,TcR和Ig对抗原的识别是在进化中广泛使用的域类型的高度进化和相当复杂的相互作用(7,10)。
IgSF域是如何进化的,它是如何适应抗原识别的特殊情况的?似乎IgSF结构域可能进化为介导相互作用,可能是在细胞表面(7)。基因重复和突变导致了大量蛋白质。 Strong等人的论文(1)解决了在编码上述抗体和TcR的特征性重排基因之前是否可以识别抗原结合V结构域的可能前体的问题。因此,在搜索原始V结构域时,人们希望找到一组具有推定的J样序列的V区序列。似乎可以合理地假设J区域在某个阶段可能通过插入转座因子而分离(11)。显然,引入许多J区域会立即显着增加可能的所有组成部分,特别是当考虑到D区域和N区域多样性时。人们可能期望V结构域与C结构域相关,并且蛋白质是与通过与其细胞质区域的相互作用发信号的能力相关的膜。
Strong等人研究了河豚(1),没有可识别的TcR或Ig,但具有紧凑的基因组,使其成为寻找可能已经成为当前V结构域的前体并且可能通过先天机制介导免疫反应的蛋白质家族的合适物种。 Strong等人使用PCR技术分离出一个名为NITR或新型免疫型受体的26个基因家族。 NITR含有两个IgSF结构域,一个跨膜区和一个细胞质区,在某些情况下含有ITIM或抑制性酪氨酸基序。 这些基序通常存在于非重排受体中,例如天然杀伤受体,但不存在于TcR或Ig中,其与含有ITAM(活化基序)的蛋白质相关。 膜近端结构域称为V / C2,因为它类似于V和C2结构域。
在人和啮齿动物白细胞表面表征的蛋白质中约有三分之一含有IgSF结构域,其中45%含有两个结构域,其中大部分也具有NITR中发现的V结构域-C2结构域 - 跨膜组织(10)。 这些包括CD2亚家族(CD2,CD48,CD58,CD84和2B4)(12)和其他如OX2,CD33,CD80和CD86(10,13)。 TcR和Ig轻链还含有具有C1结构域的氨基末端V结构域,其具有仅在抗原受体和MHC抗原中发现的序列模式。
第一次重排抗原受体的直接前体可能仍然存在于高等脊椎动物中,并且看到可以在哪种物种中发现NITR同源物将是有趣的。 类似地,脊椎动物蛋白质的同源物是否具有V-C2-跨膜排列,具有上述非重排J-样片段? 一个有趣的例子是在两栖动物和人类中发现的CTX基因家族,但它们没有显示C2结构域中的保守性组合以及NITR中看到的V结构域的广泛变异(14)。
显然,从编码原始受体的有限数量的基因开始有一个重要的步骤,其可能与NITR相关,重排TcR和Ig。 一种理论认为,这种变化发生在从无颌鱼到下颌鱼的过渡期间,这可能导致局部受伤和感染增加(15)。 制造更多抗原受体的能力可能是一个重要的选择性优势,并导致适应性免疫系统的发展。
毫无疑问,NITR蛋白家族的发现是我们对抗原受体和适应性免疫系统进化的思考的一个有趣的贡献。 发现这些蛋白质与之相互作用将是有意义的。 V域是否通过一种原始免疫来识别外来蛋白质? 或者它们是否会与其他细胞或可溶性蛋白质表面的自身蛋白质相互作用? NITR的膜近端结构域的极高保守性表明它可能以与顺式方式的保守相互作用参与另一种细胞表面蛋白,可能类似于CD3链并参与传递信号。 它们形成同源二聚体还是异二聚体? 在脊椎动物中是否存在存在已建立的适应性系统的NITR的同源物,它们是否参与免疫系统?
参考资料
- Neil Barclay.PNAS December 21, 1999 96 (26) 14672-14674.Ig-like domains: Evolution from simple interaction molecules to sophisticated antigen recognition ( https://www.pnas.org/content/96/26/14672 )
- https://prosite.expasy.org/cgi-bin/prosite/nicedoc.pl?PS50835
- https://prosite.expasy.org/PS50835
