【3.1】基于 NY-ESO-1 的癌症免疫疗法
NY-ESO-1 或纽约食管鳞状细胞癌 1 (New York esophageal squamous cell carcinoma 1)是一种众所周知的癌症睾丸抗原 (CTA, cancer-testis antigen),在多种癌症类型中重新表达。它能够引发自发的体液和细胞免疫反应,加上其受限的表达模式,使其成为癌症免疫治疗的良好候选靶点。
在这篇综述中,我们提供了正常和癌组织中 NY-ESO-1 表达和功能的背景信息。此外,已在各种癌症类型中观察到 NY-ESO-1 特异性免疫反应;然而,它们作为生物标志物的效用尚未确定。最后,我们描述了目前正在进行临床试验的针对 NY-ESO-1 的基于免疫的治疗选择。我们将重点介绍 NY-ESO-1 癌症疫苗、过继(adoptive) T 细胞疗法、和检查点抑制剂的组合治疗,并将讨论未来基于 NY-ESO-1 的免疫治疗的当前趋势。在过去的几十年里,随着免疫疗法的出现,癌症治疗发生了革命性的变化。基于免疫的干预已显示出有希望的结果,为各种癌症类型的持久临床反应提供了新的治疗途径。大多数成功的免疫疗法研究都是在液体癌症中报道的,而这些方法在实体癌症中遇到了许多挑战。复杂、动态的肿瘤微环境会调节抗肿瘤免疫反应的程度和表型,这阻碍了实体癌的有效免疫治疗。此外,许多实体瘤相关抗原不是私有的,而是可以在正常体细胞组织中找到的,导致轻微到有害的脱靶毒性。
因此,一直在努力使用各种基于免疫的方式来识别肿瘤特异性抗原。CTA 被认为是免疫治疗的良好候选靶标,因为它们的特征是在正常体细胞组织中表达受限,同时在实体上皮癌中重新表达。此外,已发现几种 CTA 可诱导自发免疫反应,NY-ESO-1 是家族成员中最具免疫原性的。因此,本综述将重点关注 NY-ESO-1,并讨论过去和当前的 NY-ESO-1 靶向免疫治疗策略。CTA 被认为是免疫治疗的良好候选靶标,因为它们的特征是在正常体细胞组织中表达受限,同时在实体上皮癌中重新表达。此外,已发现几种 CTA 可诱导自发免疫反应,NY-ESO-1 是家族成员中最具免疫原性的。因此,本综述将重点关注 NY-ESO-1,并讨论过去和当前的 NY-ESO-1 靶向免疫治疗策略。CTA 被认为是免疫治疗的良好候选靶标,因为它们的特征是在正常体细胞组织中表达受限,同时在实体上皮癌中重新表达。此外,已发现几种 CTA 可诱导自发免疫反应,NY-ESO-1 是家族成员中最具免疫原性的。因此,本综述将重点关注 NY-ESO-1,并讨论过去和当前的 NY-ESO-1 靶向免疫治疗策略。
一、癌症睾丸抗原 (CTA) Cancer-Testis Antigens
癌症-睾丸抗原形成一个由 276 个基因编码的抗原家族,包括 70 多个基因家族 ( 1 )。大约 50% 的 CTA 基因在 X 染色体上形成多基因家族,被称为 CT-X 基因 ( 2 )。这些 CTA 位于 Xq24-q28 区域 ( 3 ) 中密度最高的染色体上的特定簇中。CTA 表达主要限于睾丸生殖细胞和胎盘滋养细胞,在正常成人体细胞中没有表达或表达很低 ( 3 , 4 )。
有趣的是,已经发现几种 CTA 在多种上皮癌中重新表达。例如,使用来自基因型-组织表达项目的 RNA 测序数据集,Rooney 等人。鉴定了 60 个在正常组织中不表达而在多种肿瘤类型中异位表达的 CTA,包括黑色素瘤、头颈癌、肺癌、肝癌、胃癌和卵巢癌 ( 5 )。在癌症类型中,根据 CTA 表达的频率,肿瘤可分为富含 CTA、中等或较差。
- 富含 CTA 的肿瘤包括黑色素瘤、卵巢癌、肺癌和膀胱癌;
- CTA 中度肿瘤组包括乳腺癌、膀胱癌和前列腺癌,
- 而肾细胞癌、结直肠癌和淋巴瘤/白血病被归类为 CTA 较差肿瘤(4 , 6)。
一些报告表明,在某些肿瘤类型中可能偏好特定的 CTA 重新表达,例如 70% 的转移性黑色素瘤中的 MAGEA1-4、70% 的卵巢肿瘤中的 ACRBP 和 46% 的乳腺癌中的 NY-ESO-1肿瘤(7)。CTA 表达模式也与疾病阶段有关。例如,虽然在良性前列腺组织中没有观察到 CTA 的 SSX 家族的表达,但 23% 的转移性前列腺癌病变显示 SSX 蛋白的重新表达 ( 8 )。尽管 CTA 表达可能会在大量原发性肿瘤中增加,但这可能不会反映在单细胞水平上。例如,卵巢癌标本的显微解剖显示 NY-ESO-1 表达的相当大的肿瘤内异质性(9)。异源表达也可以在源自一个原发病变的转移灶中发现 ( 10 )。肿瘤内异质性可以部分解释某些 CTA 在肿瘤中重新表达的不同程度。此外,RNA 和蛋白质表达水平之间的差异并不少见,并且可能导致所报告的表达水平的多样性。非常需要直接比较相同样本中 RNA 和蛋白质表达水平的研究,这受到缺乏特定的市售 CTA 抗体的阻碍。
CTA 异常重新表达的潜在机制包括 DNA 去甲基化和组蛋白翻译后修饰,最近的证据也支持 miRNA 介导的调节作用 ( 2 , 11 )。有趣的是,去甲基化剂如 5-aza-2-deoxycytidine 能够诱导某些 CTA 的重新表达,特别是在肿瘤细胞中而不是在正常上皮细胞中 ( 12 – 16 )。
癌症睾丸抗原不仅在肿瘤组织中重新表达,而且还被认为是免疫原性蛋白质,因为该家族的许多成员已被证明可以在癌症患者中引发自发的细胞和体液免疫反应。发现的第一个 CTA,MAGE-A1,是通过其在黑色素瘤患者中诱导自体细胞毒性 T 淋巴细胞反应的能力而发现的 ( 17 )。从那时起,其他几种 CTA 已被鉴定为免疫原性肿瘤相关抗原 (TAA),包括 SSX-2、NY-ESO-1 以及 BAGE、GAGE 和 MAGE 家族的各种成员 ( 2 , 18 )。
尽管关于正常和肿瘤组织中 CTA 表达的数据越来越多,但它们的功能在很大程度上仍然难以捉摸。转基因小鼠模型已经揭示了几个 CTA 成员在男性生育能力中发挥关键作用 ( 7 )。少数 CTA 与细胞代谢、细胞骨架动力学、双链断裂修复、基因组完整性的维持以及精子发生过程中 mRNA 表达的调节有关。尽管越来越多的研究表明 CTA 在癌症中的重新表达,但它们在肿瘤发生中的功能作用在很大程度上尚未得到探索。最近的数据表明 CTA 在调节上皮间质转化以及肿瘤细胞存活中的作用 ( 19 )。
鉴于它们在成人体细胞组织中的表达受到很大限制,并且具有免疫原性潜力,CTA 被认为是癌症免疫治疗的良好候选靶点。迄今为止,NY-ESO-1 是最有希望的基于免疫干预的 CTA,因为它的肿瘤表达与多种恶性肿瘤的免疫反应诱导明显相关(2)。因此,本综述将重点关注 NY-ESO-1 的表达模式和生物学功能;并且还将讨论 NY-ESO-1 肿瘤免疫学和免疫疗法的一些过去和最近的发展。本综述并不旨在涵盖所有关于癌症中 NY-ESO-1 的可用文献,因为这已成为一个非常大的研究领域。因此,本综述中可能遗漏了某些研究,我们提前道歉。
二、NY-ESO-1 在正常组织中的表达
CTA 纽约食管鳞状细胞癌 1 (NY-ESO-1) 也称为癌睾丸抗原 1B (CTAG1B),由位于 X 染色体 Xq28 区域的基因CTGAG1B编码。
NY-ESO-1 是 CTA 的典型例子,它限制在生殖细胞和胎盘细胞中表达,并在肿瘤细胞中重新表达。在胚胎发育过程中,NY-ESO-1 表达最早可在 13 周时检测到,并在 22-24 周时表达最高 ( 20 )。虽然 NY-ESO-1 在精原细胞和初级精母细胞中保持表达,但其在雌性卵原细胞中的表达迅速下降 ( 3 , 20 – 22 )。已在卵巢和子宫内膜组织中检测到低水平的 NY-ESO-1 表达的 RNA,而非蛋白质,尽管其生物学相关性尚不清楚 ( 21 , 23 )。
NY-ESO-1 是一种 18-kDa 蛋白质,具有 180 个氨基酸,具有富含甘氨酸的 N 端区域和带有 Pcc-1 结构域的强疏水性 C 端区域。NY-ESO-1 蛋白已被证明与位于同一区域的另外两个 CTA 同源;LAGE-1 和 ESO3 ( 24 )。虽然 NY-ESO-1 和 LAGE-1 编码非常同源的蛋白质,在成人睾丸中的表达受限,但 ESO3 蛋白质与两者的相似性相当低,据报道在各种体细胞组织中表达 ( 25 )。筛选 cDNA 表达文库以发现 T 细胞表位表明 NY-ESO-1 可以从替代的开放阅读框产生,从而产生一个 58 个氨基酸长的蛋白质,可被肿瘤反应性 T 细胞识别 ( 26 )。
尽管对 NY-ESO-1 的生物学功能知之甚少,但可以从其结构和表达模式中推断出可能的功能。保守的 Pcc-1 结构域的存在表明它可能参与细胞周期进程和生长 ( 27 )。与黑色素瘤抗原基因 C1(CTA 的 MAGE 家族成员)共表达 ( 28 ),鉴于 MAGE 蛋白在这些细胞过程中的突出作用,进一步支持参与细胞周期进程和细胞凋亡 ( 29))。此外,其受限的表达模式表明它可能在生殖细胞自我更新或分化中发挥作用。与癌细胞中主要的细胞质表达相反,NY-ESO-1 在间充质干细胞中的核定位进一步支持了这一观点。间充质干细胞分化后,NY-ESO-1 表达下调,这表明可能在干细胞和癌细胞的细胞增殖中发挥作用 ( 30 )。此外,与分化细胞相比,神经胶质瘤癌症干细胞中 NY-ESO-1 的表达增加,同时伴有组蛋白乙酰化和 DNA 低甲基化 ( 31 )。
三、NY-ESO-1 在癌症中的表达
肿瘤表达
NY-ESO-1 在多种肿瘤类型中均有报道,包括神经母细胞瘤、骨髓瘤、转移性黑色素瘤、滑膜肉瘤、膀胱癌、食管癌、肝细胞癌、头颈癌、非小细胞肺癌、卵巢癌癌症、前列腺癌和乳腺癌 ( 21 , 32 – 46 )。在这些肿瘤类型中,NY-ESO-1的表达频率差异很大,最常见的表达肿瘤是粘液样和圆细胞脂肪肉瘤(89-100%)、神经母细胞瘤(82%)、滑膜肉瘤(80%)、黑色素瘤( 46%) 和卵巢癌 (43%) ( 37 , 47 – 51)。其他肿瘤类型显示 NY-ESO-1 的蛋白质表达范围为 20-40%。此外,许多研究报告了 RNA 表达数据,这些数据可能与免疫组织化学确定的蛋白质表达水平有很大不同。并且只有少数研究调查了 RNA 和蛋白质的表达。
需要注意的是,大多数癌症类型都表现出 NY-ESO-1 的异质表达,这可能会限制对 NY-ESO-1 靶向治疗的治疗反应。在粘液样和圆细胞脂肪肉瘤 (94%) 和滑膜肉瘤 (70%) 中报道了最同质的表达,这可能与在过继细胞免疫治疗试验中获得的有希望的结果有关 ( 47 , 49 )。
体液和细胞免疫反应
对 NY-ESO-1 的自发免疫反应的第一个证据是在一名食管癌患者中报道的 ( 52 )。使用一名食管鳞状细胞癌患者的肿瘤 mRNA,作者构建了一个肿瘤 cDNA 表达文库,并用同一名患者的血清进行了免疫筛选。利用SEREX技术( Serological analysis of recombinant cDNA expression libraries ),NY-ESO-1被鉴定为肿瘤抗原,可以诱导高滴度的IgG体液应答。
可以在多种癌症患者中检测到针对 NY-ESO-1 的体液免疫反应,包括结肠直肠癌、肺癌、乳腺癌、前列腺癌、胃癌和肝细胞癌 ( 53 – 61 )。由于这些研究大多集中在一种癌症类型上,大岛等人的研究。值得注意的是,他们对来自各种癌症类型的患者的 1,969 份样本进行了大型血清学研究 ( 62 )。作者可以检测所有癌症的血清 NY-ESO-1 自身抗体,其中食管癌频率最高 (32%),其次是肺癌 (13%)、肝细胞癌 (11%)、前列腺癌和胃癌 (10%) )、结直肠癌 (8%) 和乳腺癌 (7%)。对健康个体的分析显示没有自发的 NY-ESO-1 体液反应。
NY-ESO-1 也被证明可以诱导细胞免疫反应。在具有高滴度抗体反应的转移性黑色素瘤患者中观察到同时针对 NY-ESO-1 的体液和细胞反应的第一份报告 ( 63 )。NY-ESO-1 中的三个 HLA-A2 限制性表位被鉴定为 CD8+ 细胞毒性 T 淋巴细胞的识别位点。在一项后续研究中,同一团队通过在携带 NY-ESO-1 抗体的 11 名黑色素瘤患者中的 10 名中检测到 NY-ESO-1 特异性 CD8+ T 细胞来验证他们的发现 ( 64 )。在后来的一项研究中,他们还确定了三个 HLA-DRB40101-0103、MHC II 类表位,这些表位可被来自两名黑色素瘤患者的 CD4+ T 淋巴细胞识别(65)。此外,其他人在 NY-ESO-1 的羧基末端发现了 HLA-DRB10401 和 HLA-DP4 限制性表位,它们可以被黑色素瘤患者的 CD4+ T 淋巴细胞识别 ( 66 – 69 )。含有 HLA-DP4 限制性表位的肽也可以产生 HLA-A2 限制性 CD8+T 细胞,表明该肽可用作癌症疫苗以诱导 CD4+ 和 CD8+T 细胞反应(70)。这开辟了可用于对抗 NY-ESO-1 阳性肿瘤的免疫治疗方法的数量,这些肿瘤诱导体液免疫系统以及细胞 CD4+ 和 CD8+ T 细胞区室,单独或组合。有趣的是,在健康个体中,发现 NY-ESO-1 特异性 CD4+ T 细胞前体被调节性 T 细胞积极抑制,这表明肿瘤微环境的细胞因子环境可以支配和阻碍自然的 NY-ESO-1 抗肿瘤免疫反应。71,72)。这个概念非常重要,在设计新的基于免疫的干预措施时应予以考虑。
NY-ESO-1 作为生物标志物
已发现 NY-ESO-1 表达跨肿瘤类型与晚期疾病的几个特征相关,包括更高的分化等级、淋巴结转移和临床分期 ( 23 )。NY-ESO-1 表达作为预后生物标志物的价值仍然存在争议。虽然它的表达与某些癌症的不良临床结果有关,但在其他癌症中没有发现相关性或相互矛盾的结果。例如,在非小细胞肺癌、肝细胞癌、头颈癌、胃肠道癌、多发性骨髓瘤和恶性黑色素瘤中,NY-ESO-1 肿瘤表达与较高的复发风险、较差的治疗反应和较短的治疗时间相关。生存(73 – 83)。相反,在非霍奇金淋巴瘤中,NY-ESO-1 肿瘤表达与早期疾病相关 ( 84 )。关于卵巢癌和乳腺癌的早期报告没有显示出与预后有任何显着相关性 ( 37 , 85 , 86 )。对 NY-ESO-1 的免疫反应也在循环生物标志物的背景下进行了研究,为监测疾病进展和治疗反应提供了一种非侵入性方式。已发现 NY-ESO-1 特异性体液免疫反应的程度随着疾病的进展而增加并随着疾病的消退而减少(32 , 33 , 38 , 60 , 62 , 87 )– 89)。Esfandiary 和 Ghafouri-Fard 提供了 NY-ESO-1 在不同癌症中的表达和免疫原性的综合概述 ( 22 )。该综述发表后,新发现表明食管癌中 NY-ESO-1 抗体的检出率随着疾病分期而逐渐增加,从 I 期的 16% 到 IV 期的 42% ( 62 )。此外,在结直肠癌中,NY-ESO-1 抗体的存在最近与几个预后临床病理参数相关,包括肿瘤浸润深度、临床分期、淋巴结和远处转移(89)。NY-ESO-1 抗体随时间的变化可指示疾病消退,并可用作疾病监测的标志物,如 12 名不同肿瘤类型的患者所证明的 ( 88 )。在 NY-ESO-1 体液反应降低的五分之四的患者(两名膀胱癌、两名黑色素瘤、一名非小细胞肺癌)中,观察到肿瘤负荷和/或转移减少。同样,NY-ESO-1-T 细胞反应已被研究作为预后标志物。在转移性黑色素瘤中,循环 NY-ESO-1 特异性 T 淋巴细胞的存在与更好的预后相关,将总生存期从 6 个月提高到 21 个月 ( 90 )。
四、NY-ESO-1 定向免疫疗法
NY-ESO-1 被广泛认为是免疫治疗的良好候选靶点,并且在早期 I/II 期研究中已经取得了一些有希望的结果。NY-ESO-1 在正常组织中的有限表达结合其在肿瘤类型中的广泛表达,使 NY-ESO-1 成为具有有限脱靶毒性和广泛应用于多种癌症类型的靶标。此外,其强大的免疫原性表明有机会增强针对这种 TAA 的天然免疫反应。迄今为止,使用各种基于免疫的干预措施 ( http://www.clinicaltrials.gov )针对 NY-ESO-1 进行了 12 项活动、31 项招募和 5 项拟议的临床试验。在这篇综述中,我们将讨论正在进行的临床试验,总结在表中表11——3,并强调使用 NY-ESO-1 疫苗接种方法、过继细胞疗法或与免疫检查点抑制剂联合治疗的一些已完成试验(图 (图11)。

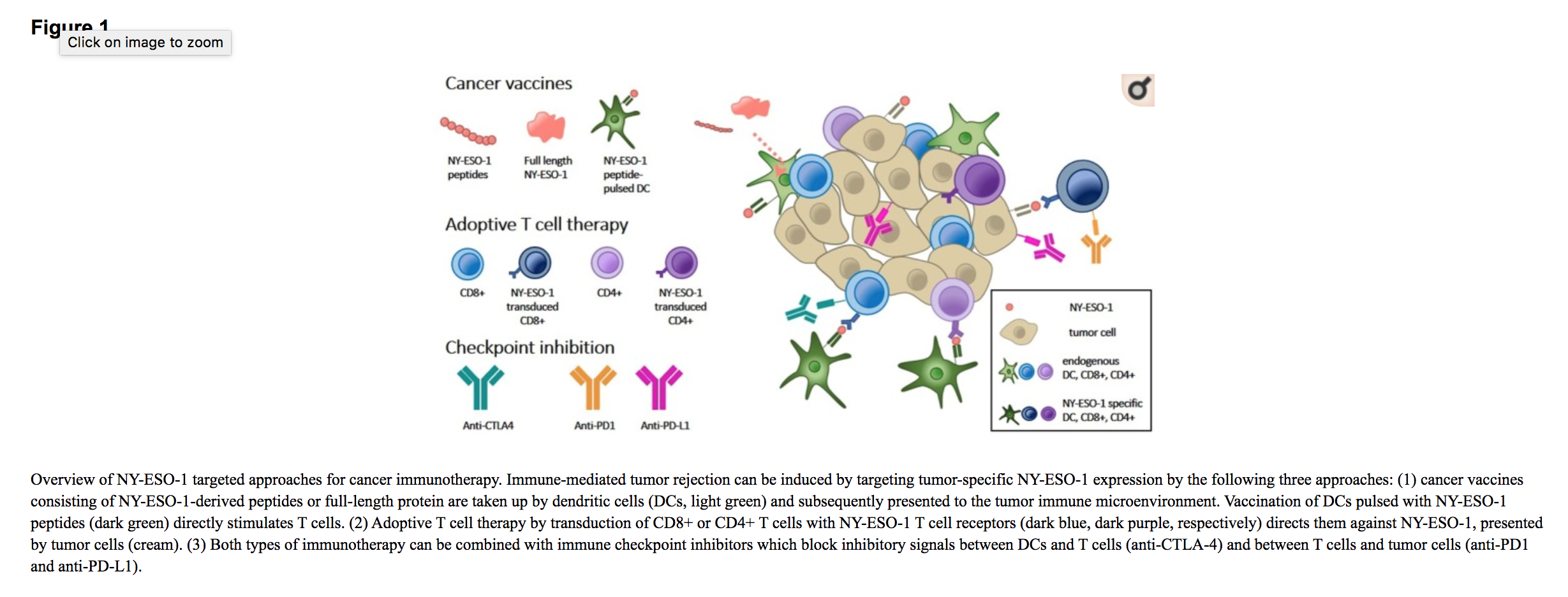
NY-ESO-1 癌症疫苗
自十多年前进行的第一次临床试验以来,目前的 NY-ESO-1 癌症疫苗试验已经有了很大的发展。在肽发现和疫苗配制方面取得了许多进展。我们现在拥有一个由单个合成肽、单个和复合重组蛋白以及各种佐剂配方组成的武器库。
- 已经对各种佐剂进行了严格的测试,以确定它们在暴露于 MHC I 类限制性肽时增强细胞毒性 CD8+ T 淋巴细胞活性的能力。这些佐剂从粒细胞/巨噬细胞集落刺激因子到 montanide-ISA-51 (Montanide)、赖氨酸和羧甲基纤维素稳定的聚肌苷酸-聚胞苷酸 (Poly-ICLC)、不完全弗氏佐剂、皂苷基佐剂 (ISCOMATRIX)胆固醇支链淀粉,91,92)。已提出多种因素的组合作为新型佐剂 (APH);由明矾、多糖和短合成先天防御调节肽 HH2 组成 ( 93 )。用这种佐剂接种 NY-ESO-1 可显着增加小鼠的体液和细胞反应并减少黑色素瘤负担。
- 另一种增强疫苗接种效力的策略是诱导 CD4+ 免疫反应,以支持 CD8+ 细胞毒性 T 淋巴细胞的启动和维持 ( 92 )。迄今为止,已在 NY-ESO-1 中鉴定了 21 个不同的表位,仅限于至少五个不同的 HLA-II 类等位基因。肽 NY-ESO-1 80–109和 NY-ESO-1 157–165分别与 CD4+ 和 CD8+ T 细胞反应相关,已被证明是最具免疫原性的。使用全长重组蛋白可以进一步增强诱导免疫反应的程度,因为 I 类和 II 类表位均可用于抗原呈递和加工。例如,单独接种重组全长 NY-ESO-1 或与 ISCOMATRIX 佐剂联合接种黑色素瘤患者会导致 NY-ESO-1 特异性抗体的强烈诱导,以及特异性 CD4+ 和 CD8+ T 的增加细胞(94)。使用更长的肽也有可能诱导更强的免疫反应。例如,接种 20-mer NY-ESO-1 91-110肽,覆盖多个表位,诱导体液和细胞 CD4+ 和 CD8+ T 细胞反应。此外,10 名癌症患者中有 3 名病情稳定 ( 95 )。
- 增强 NY-ESO-1 疫苗接种反应的另一种方法是使用初免和加强计划,包括初始疫苗接种,即初免,使用 1 种癌症疫苗,然后施用第二种疫苗,即加强。使用重组 NY-ESO-1 疫苗/重组禽痘-NY-ESO-1 疫苗初免-加强方案,已经观察到针对多种 NY-ESO-1 表位的体液和细胞反应(96 , 97)。目前临床试验中对无细胞疫苗方法的其他修改是使用细菌载体进行 NY-ESO-1 疫苗递送(NCT01967758,表表格1);1); NY-ESO-1 疫苗接种与 mTOR 抑制剂西罗莫司(NCT01536054、NCT02833506,表表格1)1) 或使用去甲基化剂地西他滨 (NCT02750995,表 表格11)。
由于树突状细胞 (DC) 是主要的抗原呈递细胞并且是 T 细胞的强激活剂,因此许多研究调查了肽脉冲 DC 作为细胞疫苗的用途 ( 98 )。该领域的进一步进展证明了将 toll 样受体 (TLR) 激动剂作为佐剂的额外好处。这背后的基本原理在于 TLR 介导的 DC 激活和 T 辅助 1 细胞反应的诱导 ( 99 )。事实上,与 TLR3 (Hiltonol)、TLR7 (咪喹莫特)、TLR7/8 (resiquimod) 或 TLR9 (CpG 7909) 激动剂联合治疗已被证明可以增强很大比例的癌症患者的体液和细胞反应 ( 100 – 102)。NY-ESO-1 脉冲 DCs 的安全性和有效性目前正在各种临床试验中进行研究(表(表格1),1),单独使用(NCT02692976、NCT01883518、NCT02334735、NCT02224599)或与 NY-ESO-1 蛋白疫苗和 TLR4 激动剂(NCT02387125)联合使用。DCs 的特异性靶向也可以使用 DEC-205/NY-ESO-1 融合蛋白 (CDX-1401) 实现,它通过 DEC-205 受体靶向 NY-ESO-1 蛋白进行 DC 内吞。这种融合蛋白目前正在临床试验中与重组 Flt3 配体 CDX-301 联合以促进 DC 发展(NCT02129075,表表格1),1),或与 IDO1 抑制剂 Epacadostat (NCT02166905,表 表格1),1),或输注肿瘤浸润淋巴细胞 (TIL)(NCT01946373,表 表3)。3)。癌症疫苗也可以通过 TLR 靶向 DC。这种方法的一个例子是使用编码 LV305 的 NY-ESO-1 慢病毒,通过 TLR3 和 TLR7 靶向 DC,在一名患有转移性、难治性滑膜肉瘤的患者中诱导强烈的细胞免疫反应,并显着疾病消退(103)。本病例报告是正在进行的 I 期临床试验的一部分(NCT02122861,表表格1)1) 研究皮内 NY-ESO-1 特异性慢病毒 DC 靶向在黑色素瘤、非小细胞肺癌、卵巢癌和肉瘤中的应用。无细胞树突状疗法的第二个例子是使用 Fc 受体介导的脂质体包封佐剂和/或药物的摄取,随后 DC 激活 ( 104 )。将 NY-ESO-1 蛋白疫苗与脂质体包裹的化疗药物多柔比星和去甲基化剂地西他滨相结合,在 67% 和 50% 的复发性上皮性卵巢癌患者中增强了特异性体液和 CD8+ 免疫反应 ( 105 )。50% 的患者 (5/10) 获得稳定的疾病,中位持续时间为 6.3 个月,一名患者获得部分缓解 (10%),持续时间为 5.8 个月。
NY-ESO-1 过继 T 细胞疗法
NY-ESO-1 癌症疫苗已被证明可引起体液和细胞反应;然而,使用这种方法几乎没有获得完整的答复。因此,针对 NY-ESO-1 的基于免疫干预的重点随着时间的推移在很大程度上转向了基因工程 T 淋巴细胞的开发。基于从疫苗研究中获得的知识,针对特定 NY-ESO-1 表位的 T 细胞已被设计并测试其根除肿瘤的能力。
使用 HLA-A2 限制性 NY-ESO-1/LAGE-1 转导 CD8+ T 细胞的过继 T 细胞疗法提高了难治性黑色素瘤和滑膜细胞肉瘤患者的临床反应率和总生存期。在第一个队列中,106 )。11 名转移性黑色素瘤患者中有 2 名表现出完全反应,1 名患者出现部分反应。六分之四的滑膜细胞肉瘤患者经历了部分反应,这是 NY-ESO-1 过继 T 细胞疗法在非黑色素瘤患者中成功的第一个证据。
在一项后续研究中,作者扩大了他们的队列,包括 9 名黑色素瘤和 12 名滑膜细胞肉瘤患者,并提供了第一组临床反应的最新信息(107)。结合两个队列,61% 的滑膜细胞肉瘤患者获得客观缓解,5 年总生存率为 14%,55% 的黑色素瘤患者总体 5 年生存率为 33%。此外,相同的过继性 T 细胞治疗导致 80% 的多发性骨髓瘤患者接近完全或完全缓解,中位无进展生存期为 19.1 个月(108)。许多临床试验目前正在研究 NY-ESO-1 转导的 CD8+ T 细胞的安全性和有效性,使用 NY-ESO-1 作为单一目标或作为多 TAA 目标的一部分。虽然目前的一些试验,总结在表中表2,2,研究 NY-ESO-1 特异性或多 TAA TCR 转导的 T 细胞在一系列晚期实体瘤(NCT03047811、NCT02457650、NCT02869217、NCT02366546)中,其他人专注于食管癌患者亚组(NCT7617) NCT03093350)、胰腺癌 (NCT03192462)、横纹肌肉瘤 (NCT02239861)、肝细胞癌 (NCT03175705)、滑膜肉瘤 (NCT03250325) 或血液癌症 (NCT02239861)。NY-ESO-1 特异性鼠 TCR 转导 T 细胞联合高剂量 IL-2 和化疗目前正在转移性癌症患者中进行评估(NCT02774291、NCT01967823,表表 2)。2)。此外,还有两项 I/II 期临床试验研究了使用亲和力增强的 T 细胞受体 (TCR) 对 NY-ESO-1 和 LAGE-1 (NY-ESO-1 c259 T细胞)在复发或晚期多发性骨髓瘤患者中(NCT01892293、NCT01352286,表表 2)。2)。同样,亲和力增强的 NY-ESO-1 转导 T 细胞的过继转移正在评估中(表(表2)2) 转移性黑色素瘤 (NCT01350401)、卵巢癌 (NCT01567891)、滑膜肉瘤 (NCT01343043)、粘液样/圆形细胞脂肪肉瘤 (NCT02992743) 和非小细胞肺癌 (NCT030269258)晚期实体癌 (NCT03159585)。此外,NY-ESO-1 或多 TAA 转导 T 细胞疗法与其他方式联合的安全性、可行性和有效性目前正在审查中,包括与去甲基化剂地西他滨(NCT03017131、NCT01333046,表表3),3), NY-ESO-1 157–165脉冲 DCs (NCT01697527, 表表3),3),转导的外周血干细胞(NCT03240861,表 表3),3),或显性负性转化生长因子-β 受体 II 转导的 TIL(NCT02650986,表 表33)。
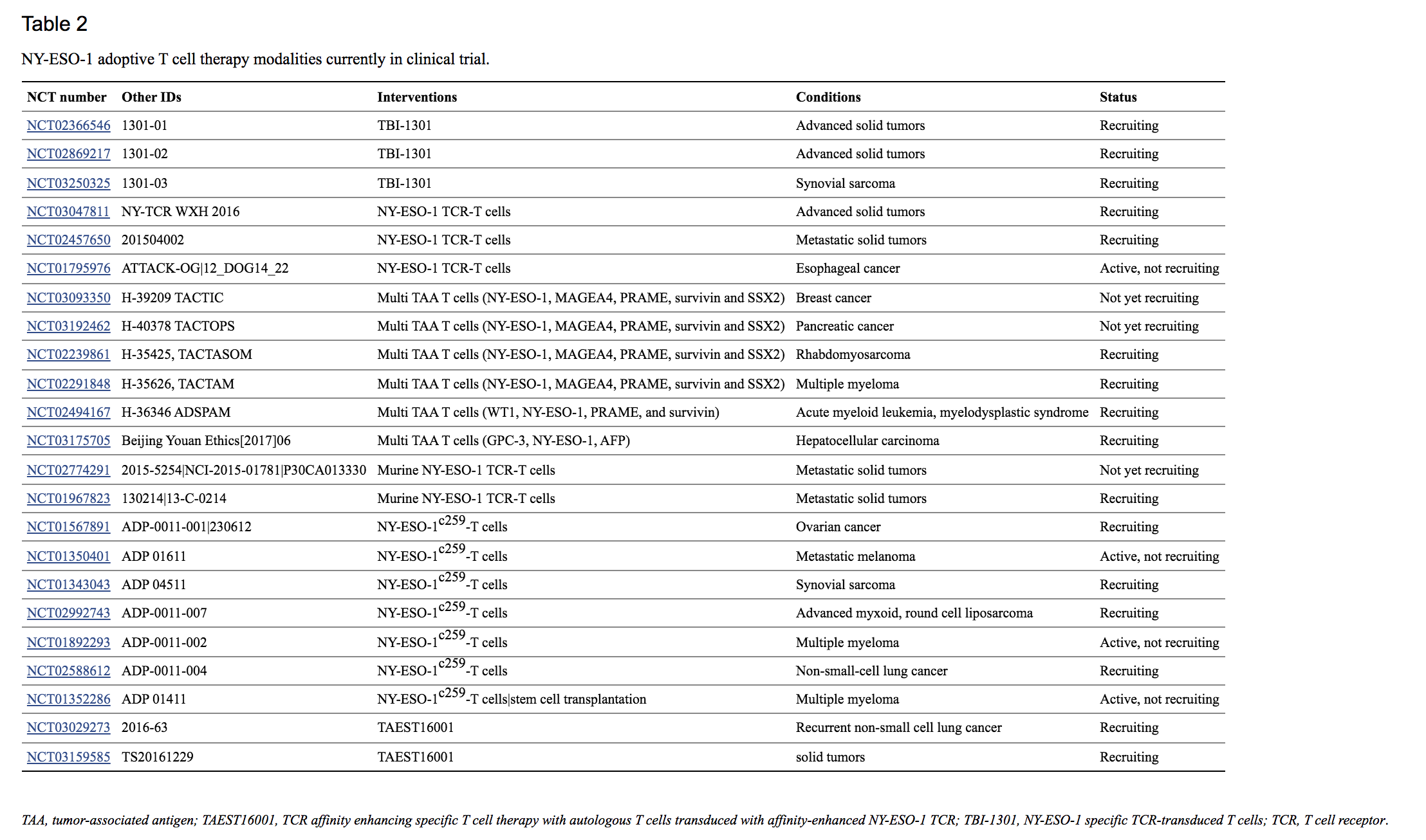
尽管大多数过继 T 细胞疗法研究都集中在细胞毒性 CD8+ T 淋巴细胞上,但再教育( re-educated)的 CD4+ T 细胞也显示出根除癌细胞的潜力。例如,一项案例研究表明,使用 HLA-DP4 限制性 NY-ESO-1 转导的 CD4+ T 细胞治疗可以诱导难治性转移性黑色素瘤的完全消退,并在 22 个月时产生持久反应(109)。然而,由于大多数癌细胞不表达 MHC II 类分子,因此基于 CD4+ 的免疫疗法本身的功效相当有限。HLA-DP 限制性 NY-ESO-1 157-170表位的呈现在体外得到增强通过使用 NY-ESO-1 和自噬分子 Atg8/LC3 的融合蛋白将抗原引导至巨自噬途径 ( 110 )。作者还报告说,通过内吞作用对 NY-ESO-1 的细胞间转移增加了抗原呈递。因此,靶向 NY-ESO-1 细胞释放和/或巨自噬的药物可以为改善 CD4+ 免疫治疗提供新的途径。MHC II 类肿瘤表达的缺乏也可以通过改造可识别 HLA I 类限制肽的 CD4+ T 细胞来解决。使用体外和体内小鼠模型,发现这种方法可以诱导癌细胞的细胞毒性和细胞因子的产生(111 – 117)。Tan等人最近的一项研究。证明 HLA-A2 限制性 NY-ESO-1 157-165转导的 CD4+ T 细胞对肽的结合亲和力高于 CD8+ T 细胞,并且能够诱导癌细胞的细胞毒性(118)。这些临床前研究结果表明,HLA 类 I 重定向的 CD4+ T 细胞可以改善当前过继性 T 细胞疗法的抗肿瘤反应。
与检查点抑制剂联合治疗
免疫反应自然受到免疫检查点分子 [细胞毒性 T 淋巴细胞相关蛋白 4 (CTLA-4)、程序性细胞死亡蛋白 1 (PD-1) 和程序性死亡配体 1 (PD-L1)] 的控制以防止过度激活导致自身免疫性疾病的免疫反应。虽然 PD-1 通路在效应阶段抑制 T 细胞活化,但 CTLA-4 通路在启动阶段的初始 T 细胞活化中发挥重要作用 ( 119 )。T 细胞上的 PD-1 与其配体 PD-L1 和程序性死亡配体 2 结合,抑制 T 细胞增殖和活化并降低 T 细胞存活率(120)。CTLA-4 是 T 细胞膜上的 CD28 同系物,通过竞争性结合将抑制 CD28:B7 结合的共刺激信号,同样导致 T 细胞增殖、分化和存活率降低 ( 121 )。CTLA-4 表达还可以增加 T 细胞的迁移率,从而减少抗原呈递细胞与 T 细胞之间的接触时间并间接抑制 T 细胞活化 ( 122 )。此外,CTLA-4 在调节性 T 细胞上持续表达,通过与 B7 的竞争性结合和/或 B7 内化的诱导,降低了抗原呈递细胞上 B7 的可用性,以形成用于 T 细胞活化的 CD28:B7 共刺激信号和生存 ( 123 , 124)。癌细胞可以劫持这些 T 细胞调节通路来抑制抗肿瘤反应。癌症免疫疗法的一种方法是通过使用特定抑制剂抑制这些检查点分子来释放自然免疫反应。对免疫检查点阻断没有反应的患者仍可能受益于癌症抗原特异性治疗的联合治疗。例如,据报道,使用 CTLA-4 检查点抑制剂 ipilimumab 治疗可在卵巢癌、前列腺癌和转移性黑色素瘤患者中诱导特异性 NY-ESO-1 体液和细胞免疫反应 ( 100 , 125 – 135)。如果伴随着 NY-ESO-1 特异性 CD8+ 细胞免疫反应,在基线时具有 NY-ESO-1 特异性体液反应的易普利姆玛治疗的黑色素瘤患者更常经历抗肿瘤反应并提高存活率 ( 131 )。
此外,从转移性黑色素瘤患者中分离出的 NY-ESO-1 特异性 CD4+ T 细胞在接受易普利姆玛治疗后能够直接裂解自体癌细胞,这表明引发 NY-ESO-1 CD4+ 细胞反应的额外临床益处(134)。总之,这些发现表明检查点抑制剂与 NY-ESO-1 靶向治疗的联合治疗可能会导致增强和更持久的临床反应。然而,这可能不适用于所有肿瘤类型。尽管 NY-ESO-1 在肉瘤中高度表达,但针对该抗原的抗体并不常见,因此质疑 NY-ESO-1 靶向治疗与检查点阻断相结合的额外临床益处 ( 136 – 138)。因此,设计了一项初步的 I 期试验,以确定 CTLA-4 阻断与 NY-ESO-1 过继 T 细胞疗法和 NY-ESO-1 疫苗在局部晚期或转移性恶性肿瘤患者中的安全性和有效性(NCT02070406,桌子表33)。
与 CTLA-4 抑制类似,免疫检查点分子 PD-1 的抑制诱导 NY-ESO-1 特异性 CD8+ 细胞毒性免疫反应 ( 139 )。支持 PD-1 抑制对 NY-ESO-1 靶向免疫疗法的潜在额外益处的进一步证据来自黑色素瘤患者的NY-ESO-1 157-165肽疫苗接种,表明 T 细胞抑制分子 PD-1 的上调,Tim -3 和 BTLA 在 NY-ESO-1 CD8+ T 细胞中 ( 140 – 143 )。PD-1 和 Tim-3 的体外阻断增加了 NY-ESO-1 157–165 CD8+ T 细胞的细胞毒性细胞增殖和细胞因子分泌( 140 , 144 )。同样,肿瘤来源的 NY-ESO-1从卵巢癌患者中分离出的92-100 个CD8+ T 细胞显示出抑制分子 PD-1 和 LAG-3 的上调,双重阻断增强了增殖和细胞因子的产生(145)。使用 PD-1 抑制剂纳武单抗联合 NY-ESO-1 157-165肽疫苗治疗转移性黑色素瘤患者显示,伊匹单抗预处理和初治患者的反应率为 25%(146)。血清学分析显示,高预处理 NY-ESO-1 CD8+ T 细胞与疾病进展相关,表明这些细胞可能表达几种抑制分子,如 Tim-3 和 LAG-3。相比之下,在对纳武单抗 PD-1 阻断有反应的非小细胞肺癌患者和无反应者之间没有观察到 NY-ESO-1 细胞免疫反应的差异 ( 147 )。支持将 PD-1 抑制剂与 NY-ESO-1 过继 T 细胞疗法一起使用的第一个实验证据来自使用肺癌异种移植小鼠模型的临床前研究(148)。在这项研究中,NY-ESO-1 转导的 T 细胞可以浸润肿瘤并减少 50% 的肿瘤生长;然而,这些细胞不能减少肿瘤负荷并显示出 PD-1、Tim-3 和 LAG-3 的上调。除了注射转导的 T 细胞外,PD-1 阻断剂还可将肿瘤负荷降低 35%。正在进行的临床试验正在探索将 NY-ESO-1 肽疫苗接种与纳武单抗相结合的安全性(NCT01176461,表表3),3),或同时使用纳武单抗和伊匹单抗(NCT01176474,表 表3)。3)。其他试验正在确定将 NY-ESO-1 转导 T 细胞疗法与纳武单抗和 NY-ESO-1 肽脉冲 DC 疫苗联合治疗晚期实体癌的安全性和可行性(NCT02775292,表表3),3),或在多发性骨髓瘤中使用派姆单抗(NCT03168438,表 表3)。3)。此外,目前正在研究将 Atezolizumab 与 NY-ESO-1 脉冲 DC 疫苗联合用于肉瘤患者的肿瘤细胞上的 PD-1 配体 PD-L1(NCT02609984,表表3),3),并与 DEC-205/NY-ESO-1 融合蛋白 (CDX-1401) 疫苗和去甲基化剂瓜地他滨联合治疗复发性卵巢癌、输卵管癌或原发性腹膜癌(NCT03206047,表) 表33)。
四、下一步去哪里?
NY-ESO-1 靶向治疗已经走过了漫长的道路,使用从肽和蛋白质疫苗接种到过继 T 细胞疗法和联合治疗方式的各种方法靶向抗原。已经获得了有希望的结果,推动了许多实体癌的新临床试验。
尽管如此,基于 NY-ESO-1 的治疗的一个紧迫问题是 NY-ESO-1 肿瘤表达的相当大的内部和内部异质性,这可能会显着限制使用 NY-ESO-1 靶向治疗根除肿瘤细胞的程度。由于包括 NY-ESO-1 在内的许多 CTA 的表达受甲基化调节,因此正在寻求的一种方法是在靶向 NY-ESO-1 之前通过去甲基化试剂增加 NY-ESO-1 的肿瘤再表达治疗。Weiser 等人的一项早期研究。证明 NY-ESO-1 表达可以通过用 DNA 去甲基化剂 5-Aza-2'-脱氧胞苷处理在体外诱导,这种效果可以通过用去乙酰酶抑制剂连续处理来增强(149)。此后,多项临床前研究表明,去甲基化不仅会增加 NY-ESO-1 在肿瘤细胞中的特异性表达,还会诱导特异性 CD8+ 免疫反应和肿瘤细胞细胞毒性;当与 NY-ESO-1 免疫疗法结合使用时,它减少了肿瘤负荷并延长了几种小鼠模型的存活率 ( 150 – 155 )。这些实验结果表明,表观遗传调节可能会增强甚至使 NY-ESO-1 过继免疫疗法在免疫原性较差的肿瘤类型中成为可能。
另一方面,尽管各种 NY-ESO-1 癌症疫苗方法已取得有希望的结果,但仍需作出一些保留。由于癌症疫苗通常基于合成肽,因此引发的问题是诱导的免疫反应是否反映或补充了针对内源性抗原表达的天然免疫反应。针对 NY-ESO-1 的自然诱导的 CD8+ 免疫反应通常针对氨基酸区域 157-165 或 157-167 内的 HLA-A2 限制性表位 ( 156 )。天然和疫苗诱导的 CD8+ 反应的比较表明,这些细胞表现出结构上保守但不同的 TCR 特征(157)。这些发现表明,用于疫苗接种的合成肽可能无法准确反映自然加工的抗原和抗肿瘤免疫反应。
另一个需要考虑的重要因素是微环境对免疫调节治疗结果的影响。癌症-免疫循环是一个众所周知的概念,并已成为免疫治疗研究的框架。癌症免疫循环描述了成功根除肿瘤细胞必须完成的各个步骤,包括癌细胞抗原的释放、癌抗原呈递、T 细胞的启动和激活、T 细胞向肿瘤的运输、T 细胞的浸润。细胞进入肿瘤,T 细胞识别癌细胞,最终杀死癌细胞(158)。因此,炎症环境的形成和免疫抑制细胞的存在会对治疗效果产生深远的影响。肿瘤细胞能够通过阻碍癌症免疫循环的每个步骤来逃避抗肿瘤免疫反应。首先,肿瘤可以通过改变肿瘤抗原的表达来逃避免疫监视。这引起了人们的担忧,即自发性肿瘤抗原免疫反应的存在可能会由于长期的免疫压力而诱导表位扩散。例如,一项 II 期研究的中期分析(NCT02609984,表表3)3) 使用 PD-L1 抑制和 NY-ESO-1 DC 靶向的组合方法显示 50% 的滑膜肉瘤和粘液样圆形细胞脂肪肉瘤患者具有特异性体液和细胞反应,但抗原表位也在 20% 的患者中扩散 ( 159)。其次,正如在炎性黑色素瘤中所证明的那样,肿瘤抗原的呈递也可以改变(160)。非炎症蛋白酶体和免疫蛋白酶体之间的活性差异已被证明会导致不同的表位库,从而阻碍 T 细胞识别和靶向肿瘤细胞的能力。
第三,免疫抑制细胞的存在或诱导会对治疗结果产生深远的影响。在晚期黑色素瘤中,显示单个 NY-ESO-1 表位可以诱导 CD4+ T 细胞反应以及刺激 T 调节细胞 ( 161 )。进一步的研究表明,这些特定的 T 调节细胞来自 CD4+ CD25- T 细胞。因此,抑制 CD4+ CD25- T 细胞向特定 T 调节细胞的外周转化可能会改善治疗结果。此外,用 NY-ESO-1/ISCOMATRIX 疫苗治疗诱导 NY-ESO-1 特异性 T 调节反应,最常见的是识别 HLA-DP4 限制的 NY-ESO-1 157-170肽(162)。在肿瘤组织中,可以观察到对 HLA-DR 限制性 NY-ESO-1 115-132肽具有特异性的 T 调节细胞。总之,这些发现表明,慢性抗原暴露可通过刺激和诱导抗原特异性 T 调节性 T 细胞来抑制循环和局部抗肿瘤免疫反应。同样,慢性乙型肝炎感染已被证明会增加肝细胞癌患者外周血和肝脏中特定 T 调节细胞的数量(163)。体外外周血单核细胞与 HBV 转染的肝癌细胞系共培养的研究表明,T 调节细胞增加以及 FoxP3 和免疫检查点 CTLA-4 的上调。有趣的是,这些 T 调节细胞不仅能够抑制 HBV 诱导的免疫反应,还能够抑制 NY-ESO-1 肿瘤抗原诱导的免疫反应。
五、总结
自发现以来,NY-ESO-1 已被研究作为基于免疫干预的抗癌目标。已经在体外、体内和临床试验中探索了几种方法。绝大多数临床试验都集中在晚期实体癌上。目前,有 12 项使用 NY-ESO-1 癌症疫苗注册的临床试验,23 项使用修饰的 T 细胞,13 项使用组合免疫疗法。随着免疫治疗领域的发展,这些方法的局限性变得越来越明显,可以通过改进当前方法或从本综述中讨论的不同角度解决这些问题来解决这些问题。探索此类新策略已经产生了目前正在临床试验中的几种新疗法
参考资料
- https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5941317/ 。 NY-ESO-1 Based Immunotherapy of Cancer: Current Perspectives 。 Published online 2018 May 1. doi: 10.3389/fimmu.2018.00947
