【8.1】靶向线粒体基因递送策略(MTS)
线粒体是细胞内的能量工厂,在细胞代谢中发挥着重要的作用,线粒体DNA(mtDNA)编码了多种蛋白,且mtDNA的突变会导致线粒体功能的紊乱,最终导致多种先天性疾病的发生。目前,靶向线粒体的基因传递系统已经被开发用来改善mtDNA 突变。然而,这些策略在线粒体基因治疗中的应用仍在探索和优化中。
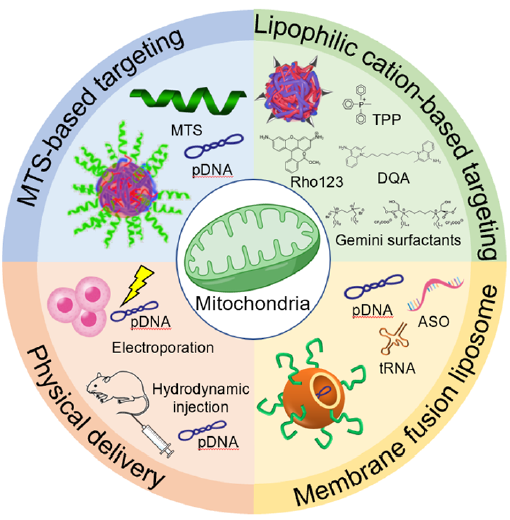
摘要图:线粒体靶向策略概述图
药物递送技术可以将多种药物(小分子药物、核酸药物、以及蛋白质)靶向到特定的细胞器、组织或细胞。靶向细胞器可以提高治疗效果,减少副作用,而线粒体因能够产生能量(ATP)、控制细胞内活性氧以及钙离子的水平、调节自噬等成为了亚细胞器中很有发展前景的靶点。此外人类的mtDNA是多拷贝、环状和双链的,包含16568个碱基对,可以编码37个基因、22种tRNA、13种对ATP合成相关的重要蛋白质以及2种rRNA。mtDNA 与核 DNA 不同,它不被组蛋白包装和保护,并且长期暴露于ROS中,因此,随着时间的推移,突变的风险会增加。mtDNA突变引起的线粒体功能障碍会导致多种遗传性疾病,如神经退行性疾病、糖尿病和癌症。除了针对人类线粒体进行基因治疗外,植物线粒体还是农业领域的重要靶细胞器,因此开发针对 mtDNA 的递送策略一直是研究热点。
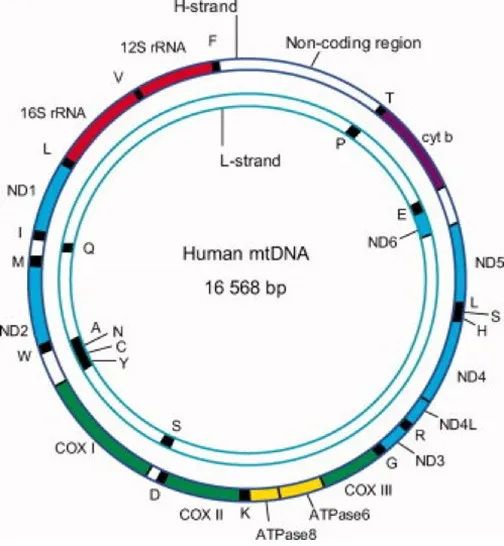
图1 mtDNA的结构
一、线粒体基因传递
如上所述,人类 mtDNA 突变与多种线粒体疾病有关,并严重破坏细胞代谢的平衡。特别是,与氧化磷酸化相关蛋白的突变会抑制 ATP 的产生。当线粒体中突变的 mtDNA 与健康的野生型 mtDNA 的比率(称为线粒体异质性)超过某个阈值时,就会发生这种线粒体功能障碍。对于线粒体疾病治疗,通过增加野生型 mtDNA或者利用基因组编辑修复突变的 mtDNA 以及消除突变的 mtDNA 来改善线粒体异质性是一种有效的策略(图2)。在线粒体基因治疗的早期阶段,健康细胞的细胞质在体外模型中可以与线粒体缺陷细胞融合,以提供完整的线粒体,通过这种方法,线粒体功能得以恢复。迄今为止,已经开发了多种用于线粒体基因治疗的策略(表1)。本文将重点介绍基于几种核酸递送的线粒体基因治疗在最近取得的成就,并根据线粒体的靶向方法进行分类。

图2 线粒体异质性导致线粒体疾病示意图
表1 靶向线粒体的核酸递送载体的总览表

二、线粒体靶向信号肽(MTS)
将质粒DNA(pDNA)引入线粒体是利用基因治疗方法提供必必需的线粒体蛋白的最为直接的方法。然而,由于线粒体外膜只对低分子量或者中等分子量的物质具有渗透性,因此需要有效的递送系统才能将质粒导入线粒体内。MTS是常用的线粒体靶向递送的工具之一。大多数线粒体蛋白在核基因组中编码然后运输到线粒体。线粒体蛋白通常在 N 端具有MTS,并表现出两亲性 α-螺旋结构,其两侧分别为阳离子和疏水残基。MTS 序列被线粒体外膜 (TOM) 复合物中带负电荷的转位酶识别,并通过桶状 TOM40 转位酶转移到线粒体膜间隙。内膜转位酶 (TIM) ,也是带负电荷的复合物,可以识别 MTS 并最终允许线粒体蛋白进入线粒体基质。这种机制已经扩展应用到线粒体靶向递送。Bennett 设计了一种基于一种功能性蛋白的 MTS 的基因传递系统,称为protofection。它由三个结构域组成:(i)蛋白质转导结构域(PTD),其序列与病毒蛋白相似,用于增强细胞摄取(ii)用于线粒体靶向的MTS结构域(iii)线粒体转录因子 A(TFAM),对mtDNA的复制至关重要。含有PTD-MTS-TFAM的野生型 mtDNA 成功转运到线粒体中,从而提高了疾病细胞系的呼吸速率和 mtDNA 拷贝数,降低了突变 mtDNA。值得注意的是,将PTD-MTS-TFAM/mtDNA 复合物静脉注射到小鼠尾静脉可提高大脑和骨骼肌线粒体中 NADH 泛醌氧化还原酶驱动的呼吸速率。
他们还报道了由 pDNA 和 MTS 缀合的肽制备的线粒体靶向胶束(图3)。由含有部分细胞色素 c 氧化酶(cytocox) 亚基 IV 序列的 MTS 与赖氨酸-组氨酸 (KH) 重复序列结合,通过静电相互作用促进纳米颗粒 (NP) 与 pDNA 形成。
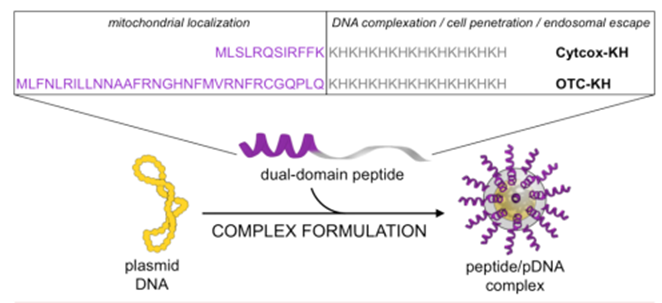
3 peptide/pDNA复合物示意图
基于 MTS 的线粒体靶向策略扩展到病毒基因传递。2012年John Guy等人开发了一种 MTS 结合的腺相关病毒 (AAV),其中装载有编码 NADH 泛醌氧化还原酶亚基 4(pND4) 的 pDNA,AAV载体的MTS递送增强了pND4在线粒体中的积累,导致视力丧失小鼠的视神经萎缩恢复。
三、基于亲脂性阳离子的线粒体靶向
亲脂性阳离子,如 TPP,作为配体分子是线粒体靶向基因递送以及小药物递送的可靠方法,因为亲脂性阳离子可以增强递送载体和线粒体之间的静电相互作用。Faria 及其同事通过酰胺偶联将 TPP 与高分子量试剂聚(乙二醇)-聚(乙烯亚胺)(PEG-PEI)结合,这增强了 pDNA 内化到线粒体和细胞吸收。最后,PEG-PEI-TPPNPs 显着增强了报告基因和 ND4 蛋白的表达(图4)。
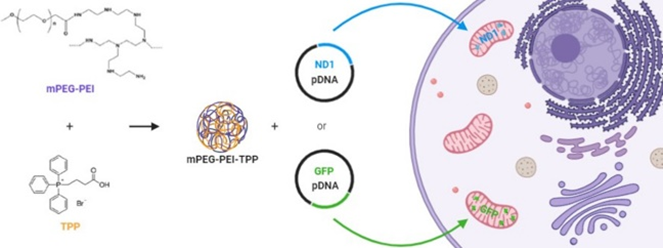
图4 PEG-PEI-TPP示意图
BeiraInterior大学的一个课题组开发了罗丹明 123(Rho123) 修饰的基因传递载体,用于线粒体靶向。为了构建载有 pDNA 的 Rho123 标记的 NP,作者使用了一种共沉淀方法,该方法是在碳酸根存在的条件下由 Ca 2+和 pDNA之间的相互作用诱导的。有趣的是,纳米粒子的大小是通过加入纤维素或明胶作为稳定剂来控制的,通过共聚焦激光显微镜观察线粒体中的荧光强度,可以发现NPs主要在线粒体中积累并有效地促进了报告基因的表达。
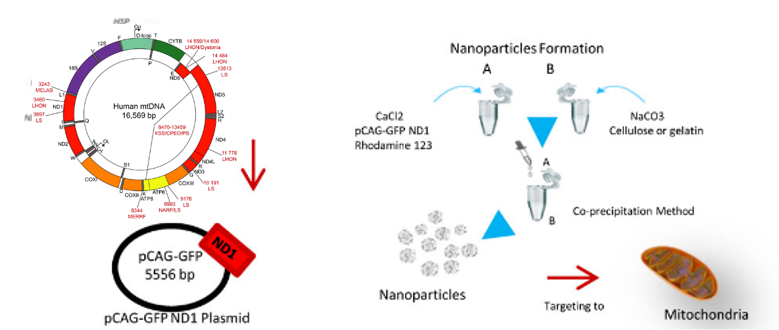
图5 Rho123修饰的基因传递载体示意图
基于Dequalinium(DQA: 1,1'-(1,10- decamethylene- bis- [aminoquinaldinium])-chloride) 与pDNA 复合的NPs被称为DQAsomes,也被认为有前途的线粒体靶向载体。DQA 是 FDA 批准的阳离子脂质,可以通过静电相互作用与 pDNA 形成脂质体样结构。载有编码 GFP 的 pDNA 的DQAsomes 在细胞中可以诱导线粒体基因表达。然而,线粒体中DQAsomes 的转染效率仅限于 5%。为了解决这个问题,Choi 和同事将 DQA 与其他脂质体——1,2-二油酰-3-三甲基铵-丙烷 (DOTAP)和 1,2-二油酰-sn-甘油-3-磷酸乙醇胺 (DOPE) 混合,以进一步改善DQAsomes 的功能。DQA 和DOTAP/DOPE 的联合使用提高了细胞摄取和内体逃逸能力,进一步增强了线粒体基因表达。
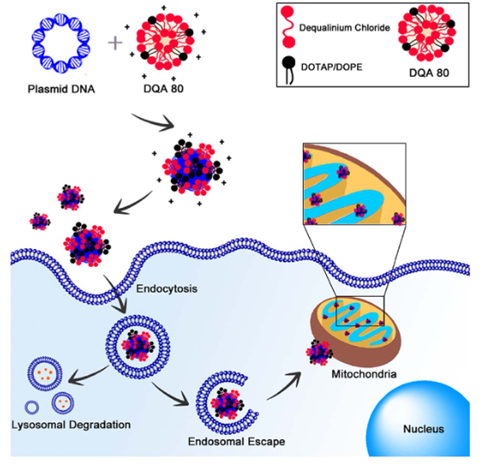
图6 DQA-DOTAP/DOPE 线粒体靶向载体示意图
四、基于脂质的线粒体融合递送
另一种基于脂质的递送载体是MITO-Porter,由 DOPE、鞘磷脂 (SM) 和细胞穿透肽 (CPP)、八个精氨酸 (R8) 组成。MITO-Porter可以通过 DOPE 诱导的膜融合机制将装载的药物输送到线粒体,R8可以增加细胞吸收的速度。这种膜融合系统能够将各种药物输送到线粒体。为了提高使用MITO-Porter 将外源 pDNA 转染到线粒体中的效率,Yamada 等人使用另一种 CPP,即具有赖氨酸-亮氨酸-丙氨酸重复序列的 KALA 肽往细胞内运输药物。KALA 修饰的MITO-Porter 构建体(KALA-MITO-Porter) 的转染效率比 R8 修饰的 MITO-Porter 构建体高约 10倍;然而,不幸的是用 KALA 替代 R8 增加了细胞毒性,这很可能是由于 KALA 肽诱导的线粒体膜的不稳定。于是作者通过整合额外的线粒体 RNA 适体 (RP) 配体,进一步增强了KALA-MITO-Porter 的功能,以增强细胞摄取和线粒体靶向活性。RP 修饰显着提高了载有 pDNA 的KALA-MITO-Porter 构建体进入线粒体的转染效率。此外,RP/KALA-MITO-Porter 的给药并未改变小鼠血清中的生物标志物水平,表明其在体内应用中具有良好的生物相容性(图7)。
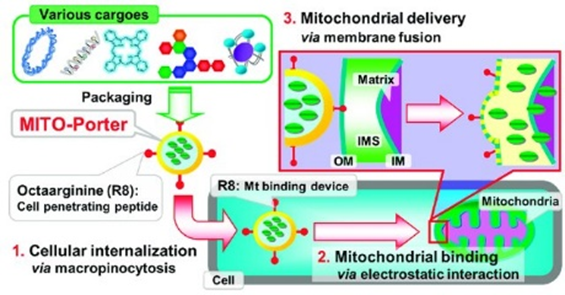
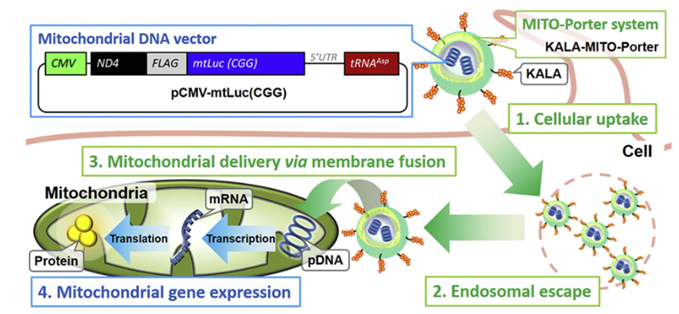
图7 两种MTIO-Porter(R8和KALA)工作示意图
五、线粒体基因组编辑
线粒体基因治疗的另一个方向是 mtDNA 的基因组编辑,这可以进一步加深了解线粒体内生物合成和线粒体疾病发生的潜在机制。通过对限制性内切酶 (RE)、锌指核酸酶 (ZFN) 和转录激活因子样效应核酸酶 (TALEN)等进行核基因组编辑的大量研究有助于 mtDNA 编辑技术的发展。由于这些编辑体系是基于编辑的蛋白质与DNA 之间的相互作用,因此将MTS 与其结合可以加速导入线粒体。此外,2020年Mok 等人通过使用 MTS 和 TALE 缀合的双链 DNA 脱氨酶毒素 A(DddA) 能够高特异性和准确性的将 mtDNA 中的 C·G 转化为 T·A,可以用于构建具有特定序列的 mtDNA 突变的疾病模型。此外,该技术还扩展到小鼠和植物线粒体以及叶绿体基因组编辑。
六、 总结
线粒体对细胞内的生命活动正常运行有至关重要的作用,比如脂肪酸氧化、TCA循环、自噬等,因此DNA紊乱会导致许多疾病的发生发展。目前常见的修复体内突变的mtDNA的方法有三种:基于亲脂性阳离子的线粒体靶向;基于脂质的线粒体融合递送;基于线粒体靶向信号肽(MTS)的靶向技术(图8)。此外基因组编辑的方法在体内被证明是有效的,但可能会在宿主细胞中引起细胞毒性。因此将基因直接传递到线粒体应该更安全,想要实现安全的体内递送,载体必须具有除靶向线粒体外的其他基本能力,包括高胶体稳定性以保护载有的基因免受核酸酶降解。另外需要考虑的是,由于线粒体疾病引起的线粒体功能障碍可以影响到各个器官,因此递送载体必须能够将载有的基因传递到多个器官,并且不会被肝脏或脾脏迅速清除。为了能够有较好的生物分布,基于MTS 的 NPs 可能不太适合,因为由阳离子和疏水残基组成的 MTSs 可能与生物大分子发生相互作用并在生理环境中形成聚集体。因此,需要设计更加复杂多样的NP 组件设计来优化生物分布。总之,未来的发展应该主要集中在载体设计上,以克服体内线粒体递送中的障碍,当最终克服这些限制时,线粒体生物合成的机制将会被深刻了解,并有可能开发针对遗传性线粒体疾病的治疗方法。
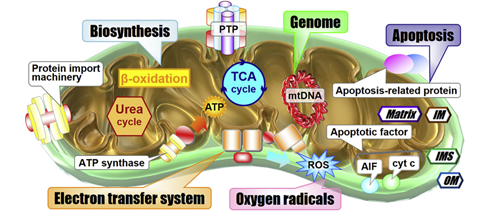
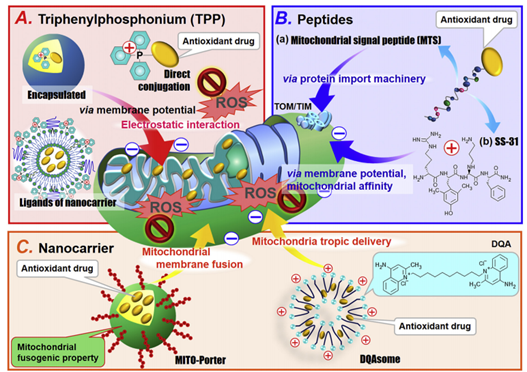
图8 线粒体与多种通路密切相关;常见的靶向线粒体传递系统
参考资料
- https://mp.weixin.qq.com/s/tu9HfxmwBsfM5JHAYKM-dQ 1.Nissanka, N., 2018. Mitochondrial DNAdamage and reactive oxygen species in neurodegenerative disease. FEBS Letters592, 728–742.
-
Chuah, J.-A., Matsugami, A., Hayashi, F.,Numata, K., 2016. Self-Assembled Peptide-Based System forMitochondrial-Targeted Gene Delivery: Functional and Structural Insights.Biomacromolecules 17, 3547–3557.
-
Yu H, Lewin AS, Guy J. Gene delivery tomitochondria by targeting modified adenoassociated virus suppresses Leber’shereditary optic neuropathy in a mouse model. Proc Natl Acad Sci U S A. 2012.
-
Physicochemical characterization andtargeting performance of triphenylphosphonium nano-polyplexes,Journalof Molecular Liquids, 2020.
-
Mitochondrial Gene Therapy: Advances inMitochondrial Gene Cloning, Plasmid Production, and Nanosystems Targeted toMitochondria,2017.
-
Choi, J.S., 2017. Functional nanosome forenhanced mitochondria-targeted gene delivery and expression. Mitochondrion 37, 27–40.
-
Yamada, Y., 2018. Mitochondrial transgeneexpression via an artificial mitochondrial DNA vector in cells from a patientwith a mitochondrial disease. Journal of Controlled 8.Mok, B.Y., 2020. A bacterial cytidinedeaminase toxin enables CRISPR-free mitochondrial base editing. Nature 583,631–637.
-
Power of mitochondrial drug deliverysystems to produce innovative nanomedicines
-
Yoshinaga, N., 2022. Rational Designs atthe Forefront of Mitochondria-Targeted Gene Delivery: Recent Progress andFuture Perspectives. ACS Biomaterials Science & Engineering.
