【3.2.1】B型DNA
“…It has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material. The structure itself suggested that each strand could separate and act as a template for a new strand, therefore doubling the amount of DNA, yet keeping the genetic information, in the form of the original sequence, intact. "
Watson和Crick最早提出来的DNA双螺旋结构实际上属于所谓的B型DNA(B-DNA)。
1953年,Watson和Crick以非凡的洞察力,以立体化学上的最适构象建立了一个与DNA的X射线衍射数据相符的DNA分子结构模型。他们与Wilkins和Franklin于1953年4月25日,在Nature上提出了DNA双螺旋模型和实验证据,解释了当时所知道的DNA分子的一切理化性质,并将DNA分子的结构特性与其携带和传递遗传信息的功能联系起来。这是一个能够在分子水平上阐述遗传的基本特征的DNA二级结构。
一、Watson和Crick提出的DNA双螺旋模型的要点如下:
(1)主链
DNA的密度表明DNA螺旋由两条绕同一轴心旋转的多聚核苷酸链组成,脱氧核糖通过3′,5-磷酸二酯键相连形成主链,两条主链反向平行,并呈右手螺旋。主链处于螺旋的外侧,所谓双螺旋就是针对两条主链的形状而言的。
(2)碱基位置
螺旋恒定的直径表明每条链的碱基局限于螺旋内部,碱基的分子平面与纤维轴垂直。碱基通过糖苷键与主链糖基相连。同一平面的碱基在两条主链间形成碱基对。但螺旋周期内的各碱基对平面的取向均不同。碱基对具有二次旋转对称性的特征,即使碱基旋转180°也不影响双螺旋的对称性。即双螺旋结构在满足两条链碱基互补的前提下,DNA的一级结构并不受限制。这一特征能很好地阐明DNA作为遗传信息载体在生物界的普遍意义。

(3)碱基配对
一条链的碱基与另一条链的碱基通过氢键联系起来形成碱基对。为了形成氢键,嘌呤总是与嘧啶配对,因为嘌呤与嘌呤配对使螺旋直径太大,而嘧啶与嘧啶配对则使螺旋直径太小。嘌呤与嘧啶配对形成专一碱基对:A和T,G和C。在碱基上形成氢键的位置为嘌呤的1位对嘧啶的1位;嘌呤的6位对嘧啶的6位。A和T之间可以形成2个氢键,G和C之间可以形成3个氢键。

(4)大沟和小沟
大沟(major groove)和小沟(minor groove)分别指双螺旋表面凹下去的较大沟槽和较小沟槽。从双螺旋中心道两条主链的连线将DNA的平面划分为两个不等的扇形,一个大于180°,一个小于180°,分别对应于大沟和小沟。在大沟和小沟内的碱基对中的N和O原子朝向分子表面。这些特性对于DNA双螺旋结构中遗传信息的识别是非常重要的,因为只有在沟内,非组蛋白才能识别出不同的碱基顺序。在双螺旋结构的表面是没有特异差别的磷酸和脱氧核糖骨架,并不携带任何遗传信息。
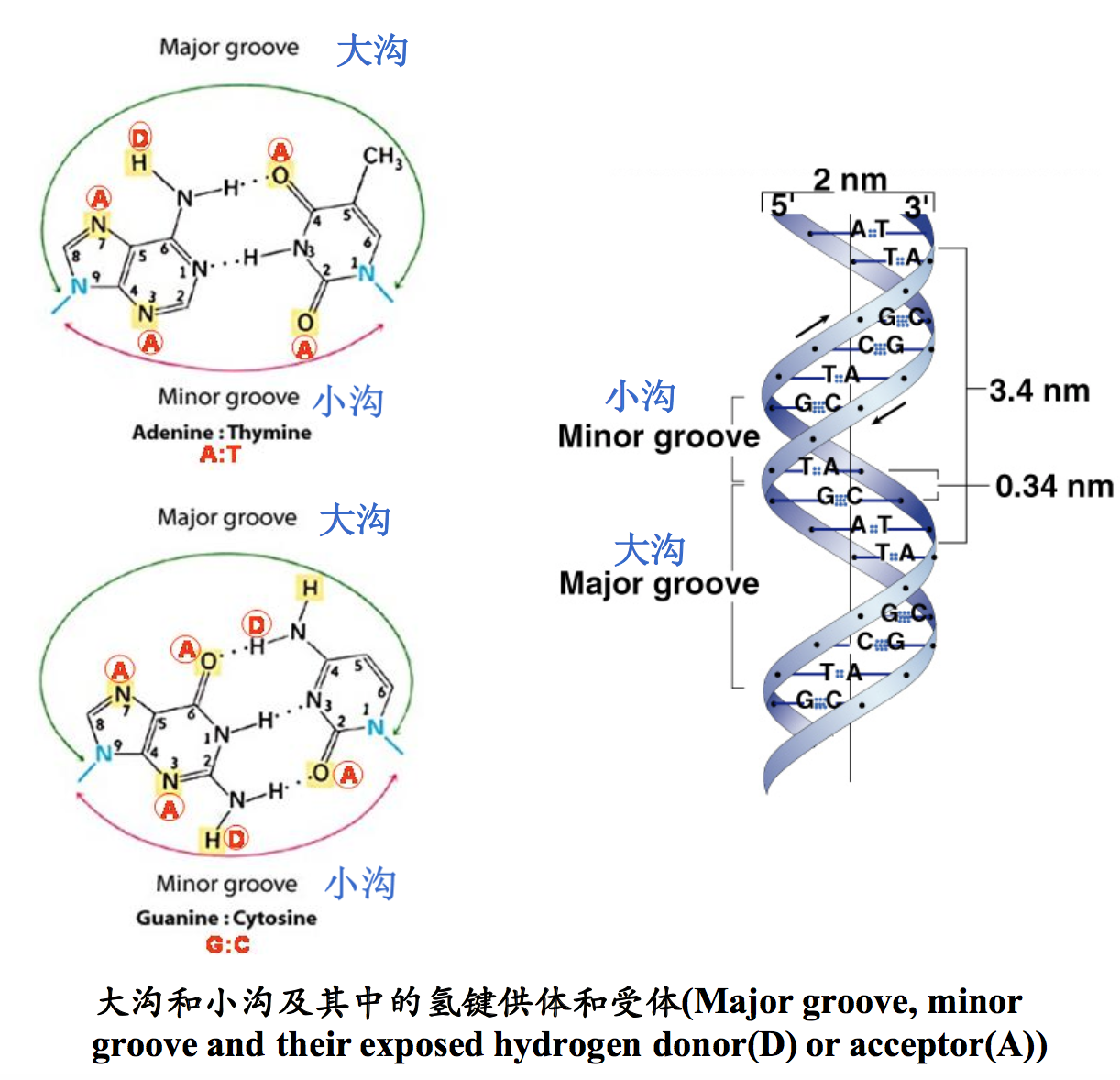
(5)螺旋参数
X射线衍射数据表明DNA具有规则的螺旋结构,螺旋直径为2nm,每一圈螺旋的高度为3.4nm,每一圈螺旋由10bp组成,相邻核苷酸之间呈36°角,距离为0.34nm。碱基倾角-2°,碱基平面基本上与螺旋轴垂直,螺旋轴穿过碱基对,
(6)维持DNA双螺旋结构的因素
影响DNA双螺旋结构的因素有以下四种力:氢键、碱基堆集力、带负电荷的磷酸基团的静电排斥力和碱基的分子内能。前二者(互补碱基的氢键结合力和相邻碱基的堆集力)有利于DNA双螺旋构象的维持,而后二者(磷酸基的静电斥力和碱基分子的内能)则不利于DNA双螺旋构象的维持。一种DNA分子的结构状态是这四种因素综合竞争的结果。
二、支持DNA双螺旋结构的关键证据 (Key evidence of DNA’s double-helix)
- Chargaff规则(Erwin Chargaff had the pairing data, but didn’t understand its implications)
- 富兰克林对DNA纤维清晰的X射线衍射照片(Rosalind Franklin’s X-ray fiber diffraction data was crucial)
- 碱基互变异构(Base tantomerism)

参考资料
- 南京大学 杨荣武老师 《结构生物学》课件
