【2.1.2.2】Hoogsteen base pair
Hoogsteen碱基对是核酸碱基配对(例如A·T对)的变体。以这种方式,两个核碱基(每条链上一个)可以通过主沟中的氢键结合在一起。Hoogsteen碱基对应用嘌呤碱基的N7位置(作为氢键受体)和C6氨基(作为供体),结合嘧啶碱基的沃森-克里克(N3-C4)面。
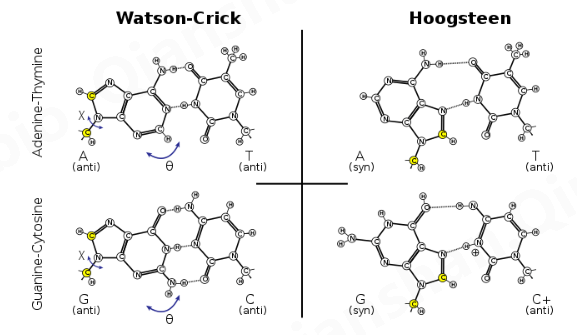
Chemical structures for Watson–Crick and Hoogsteen A•T and G•C+ base pairs. The Hoogsteen geometry can be achieved by purine rotation around the glycosidic bond (χ) and base-flipping (θ), affecting simultaneously C8 and C1′ (yellow)
一、历史
詹姆斯·沃森 (James Watson)和弗朗西斯·克里克 (Francis Crick)发表 DNA 双螺旋模型十年后, Karst Hoogsteen报道了一种复合物的晶体结构,其中 A 和 T 的类似物形成了一个碱基对,该碱基对具有不同的几何形状沃森和克里克描述过。类似地,G•C 对也可以出现另一种碱基配对几何结构。Hoogsteen 指出,如果 DNA 中存在替代的氢键模式,那么双螺旋就必须呈现完全不同的形状。Hoogsteen 碱基对可在替代结构中观察到,例如DNA 和 RNA 中形成的 四链G-四链体结构。
二、化学性质
Hoogsteen 对具有与Watson-Crick 碱基对截然不同的特性。与常规几何结构相比,两个糖苷键之间的角度(A• T 对中约为 80°)更大,而 C1 ' –C1 ‘距离(约 860 pm 或 8.6 Å)更小。在某些情况下,一个碱基相对于另一个碱基旋转 180°, 称为反向 Hoogsteen 碱基对。
在一些 DNA 序列中,尤其是 CA 和 TA 二核苷酸中,Hoogsteen 碱基对作为瞬时实体存在,与标准 Watson-Crick 碱基对处于热平衡状态。瞬态物质的检测需要使用应用于大分子的核磁共振弛豫色散光谱。[1]
在蛋白质-DNA 复合物中观察到了 Hoogsteen 碱基对。一些蛋白质已经进化到仅识别一种碱基对类型,并利用分子间相互作用来改变两种几何结构之间的平衡。
DNA 具有许多特征,使其能够被蛋白质进行序列特异性识别。这种识别最初被认为主要涉及氨基酸侧链和碱基之间的特定氢键相互作用。但很快我们就发现,没有可识别的一对一对应关系——也就是说,没有简单的代码可供阅读。部分问题在于 DNA 可能会发生构象变化,从而扭曲经典的双螺旋结构。由此产生的变异改变了 DNA 碱基向蛋白质分子的呈现,从而影响识别机制。
由于双螺旋中的扭曲本身取决于碱基序列,因此蛋白质能够以类似于识别其他蛋白质和小配体分子的方式识别DNA,即通过几何形状(而不是特定序列)。例如,A和T碱基的延伸可以导致DNA小沟变窄(双螺旋中两个沟中较窄的一个),从而增强局部负静电势,从而为带正电的精氨酸氨基创建结合位点。蛋白质上的酸残基。
三、 三重结构
DNA 三螺旋结构中的碱基三联体。 这种非沃森-克里克碱基配对允许第三链缠绕在以沃森-克里克模式组装的双链体上,并形成三链螺旋,例如(poly(dA)•2poly(dT))和(聚(rG)•2聚(rC))。在转移RNA的三维结构中也可以看到,如T54•A58和U8•A14。
三螺旋碱基配对 Watson-Crick 碱基对用“•”、“-”或“.”表示。(例如:A•T,或poly(rC)•2poly(rC))。
Hoogsteen三链 DNA碱基对用“”或“:”表示(例如:C•GC+、T•AT、C•GG 或 T•A*A)。
四、 四联体结构 Quadruplex structures
Hoogsteen 对还允许形成富含 G 的单链 DNA 和 RNA 的二级结构,称为G 四联体(G4-DNA 和 G4-RNA)。存在 G4 的体外和体内形成的证据。基因组 G4 被认为可以调节基因转录,并在 RNA 水平上通过核糖体功能的空间抑制来抑制蛋白质合成。它需要四个三联体 G,由短间隔物隔开。这允许组装由 Hoogsteen 键合鸟嘌呤分子的堆叠缔合组成的平面四重体。[8]
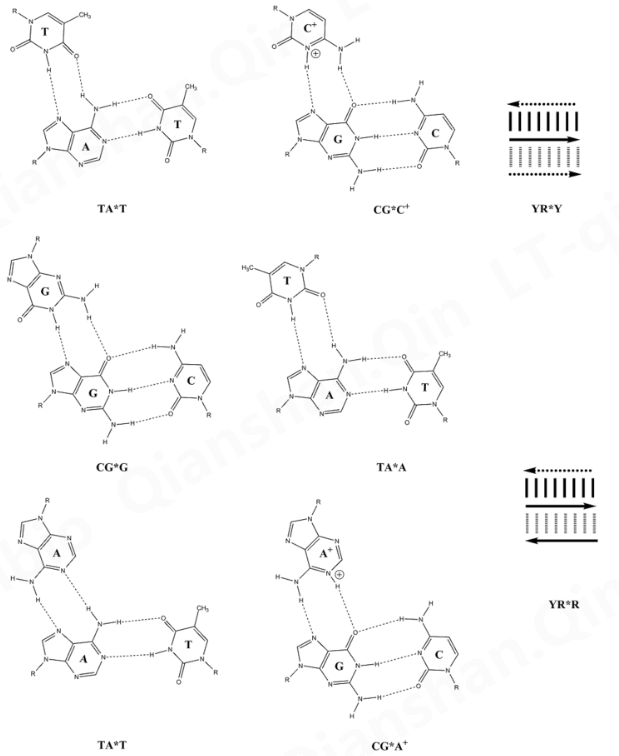
Base triads in a DNA triple helix structure.
参考资料
