【1.2.5】免疫细胞膜分子--主要组织相容性抗原(MHC/HLA)
主要组织相容性复合体(major histocompatibility complex, MHC)是一组决 定移植组织是否相容、与免疫应答密切相关、紧密连锁的基因群。哺乳动物都 有 MHC。小鼠的 MHC 称为 H-2 基因复合体;人的 MHC 称为人类白细胞抗原 (human leukocyte antigen, HLA)基因复合体。主要组织相容性抗原系统广泛分 布于人或动物有核细胞表面,其化学成分是脂蛋白或糖蛋白,在生理条件下, 此类抗原的主要功能是结合、提呈抗原肽,启动适应性免疫应答。
MHC的首次描述是由英国免疫学家Peter Gorer于1936年提出的。[6] 首先在近交小鼠品系中鉴定MHC基因。 Clarence Little在不同菌株中移植肿瘤,并根据宿主与供体的菌株发现移植肿瘤的排斥反应[7]。 乔治·斯内尔选择性繁殖了两种小鼠品系,获得了与其中一种祖先菌株几乎相同的新菌株,但在组织相容性方面差别很大 - 即移植后的组织相容性 - 因此确定了MHC基因座。[8] 在这项工作中,Snell与Baruj Benacerraf和Jean Dausset一起获得了1980年诺贝尔生理学或医学奖。
人MHC I类和II类也称为人白细胞抗原(HLA)。 为了澄清用法,一些生物医学文献使用HLA特异性地指代HLA蛋白质分子并保留编码该分子的基因组区域的MHC,但这不是一致的惯例。
一、人类 MHC 结构及其遗传性
MHC 结构十分复杂,显示多基因性和多态性。多基因性指基因复合体由多 个紧密相邻的基因座位组成,编码产物具有相同或相似的功能。组成 MHC 的 各种基因传统上分为I类、II类和III类。由于大量非经典 MHC 基因的发现, 近来倾向于以两种类型加以概括:
- 经典的I类基因和经典的II类基因,它 们的产物具有抗原提呈功能,显示极为丰富的多态性,直接参与 T 细胞的激活 和分化,参与调控适应性免疫应答;
- 免疫功能相关基因,包括传统的III类 基因,以及新近确认的多种基因,它们或参与调控固有免疫应答,或参与抗原 加工,不显示或仅显示有限的多态性。
1.1 经典的 HLAI类及II类基因
HLA 基因复合体位于人第 6 号染色体短臂 6p21.31,全长 3600kb,共有 224 个基因座位,其中 128 个为功能性基因(有产物表达),96 个为假基因。 经典的 HLAI类基因集中在远离着丝点的一端,包括 B、C、A 三个座位,其 产物称 HLAI类分子。实际上,I类基因仅编码I类分子异二聚体中的重链, 轻链为 β2 微球蛋白(β2m),编码基因位于第 15 号染色体。HLAII类基因在 复合体中位于近着丝点一端,结构较为复杂,由 DP、DQ 和 DR 三个亚区组 成。每一亚区又包括两个或两个以上的功能基因座位,分别编码分子量相近的 α 链和 β 链,形成 DRα-DRβ、DQα-DQβ 和 DPα-DPβ 三种异二聚体。

研究最多的HLA基因是9种经典MHC基因:HLA-A,HLA-B,HLA-C,HLA-DPA1,HLA-DPB1,HLA-DQA1,HLA-DQB1,HLA-DRA和HLA-DRB1。 在人类中,MHC基因簇分为三个区域:I,II和III类。 A,B和C基因属于MHC I类,而6个D基因属于II类。
1.2 免疫功能相关基因
免疫功能相关基因(immune function-related genes)分布于 HLA 复合体的 I类和II类基因区内以及III类基因区。通常不显示或仅显示有限的多态性。基 因产物除非经典性I类分子和 MIC 分子外,一般不能和抗原肽形成复合物,故 不参与抗原提呈,但它们在固有免疫应答和免疫调节中发挥重要作用。
1.2.1 血清补体成份编码基因
此类基因属经典的 HLA III类基因,在 HLA 复合体中它们位于中部(图 6- 1)。所表达的产物为 C4B、C4A、Bf 和 C2 等补体成分。
1.2.2 抗原加工提呈相关基因
蛋白酶体 β 亚单位(proteasome subunit beta type,PSMB)基因
包括 PSMB8 和 PSMB9,编码胞质溶胶中蛋白酶体的 β 亚单位。
抗原加工相关转运体(transporter associated with antigen processing, TAP)基因
产物为内质网膜上的一个异二聚体分子,分别由 TAP1 和 TAP2 两个座位的基因编码。TAP 参与对内源性抗原肽的转运,使其从胞质溶胶进入 内质网腔,并与 MHCI类分子结合。
HLA-DM 基因
包括 DMA 和 DMB 座位,其产物参与 APC 对外源性抗 原的加工提呈,帮助溶酶体中的抗原片段进入 MHCII类分子的抗原结合槽。
HLA-DO 基因
包括 DOA 和 DOB 两个座位,分别编码 DO 分子的 α 链 和 β 链。DO 分子是 DM 功能的负向调节蛋白。
TAP 相关蛋白基因
其产物称 tapasin,即 TAP 相关蛋白 (TAP- associated protein),对 I 类分子在内质网中的装配起作用,参与内源性抗原的 加工和提呈。上述抗原加工提呈相关基因全部坐落在 HLA 系统的II类基因区。
1.2.3 非经典I类基因
非经典I类基因又称 HLAIb,即 b 型I类基因,包括 HLA-E、HLA-F、 HLA-G 等。与之相对应,经典的I类基因则称为 a 型I类基因,简称 HLAI a。HLAIb 中目前研究得比较多的有下列两类基因。
HLA-E
产物由重链(α 链)和 β2m 组成,已检出 11 种等位基因。 HLA-E 分子可表达于各种组织细胞,在羊膜和滋养层细胞表面高表达。其抗原 结合槽具有高度的疏水性,能结合一种结构十分保守的 9 肽,即 HLA-Ia 和一 些 HLA-G 分子的信号肽。功能上已确定,HLA-E 分子是表达于 NK 细胞和部 分 CTL 表面的 C 型凝集素受体超家族中 CD94/NKG2 家族的专一性配体。由于 其与抑制性受体结合的亲和力明显高于与活化性受体结合的亲和力,造成生理 条件下,NK 细胞处于抑制状态。这在病毒逃避免疫监视和母胎耐受形成中可 能起十分重要的作用。
HLA-G
结构和经典性 HLA-A2 基因高度同源,由重链和 β2m 组成。 HLA-G 分子主要分布于母胎界面绒毛外滋养层细胞,相应的受体为杀伤细胞免 疫球蛋白样受体(KIR)家族成员,因而 HLA-G 在母胎耐受中发挥功能。
1.2.4 炎症相关基因
在 HLA III类基因区靠I类基因一侧,新近检出多个免疫功能相关基因 (参见图 6-1 左上方),多数和炎症反应有关,分属以下四个家族:
- 肿瘤坏死因子基因家族 包括 TNF、LTA 和 LTB 三个座位。其产物参 与炎症、抗病毒和抗肿瘤免疫应答。
- 转录调节基因或类转录因子基因家族 包括类 I κB(I κBL)基因,可 参与调节 NF κB 的活性。
- MHCI类链相关分子(MHC classIchain-related, MIC)基因家族 包 括 MICA 和 MICB 基因,其中 MICA 座位已检测到 84 个等位基因。
- 热休克蛋白基因家族 包括 HSP70 基因,其产物参与炎症和应激反 应,并作为分子伴侣在内源性抗原的加工提呈中发挥作用。
1.2.5 备注
灰色短尾负鼠(Monodelphis domestica)中的相同标记,有袋动物,跨度3.95 Mb,产生114个基因,87个与人类共享。[11] 有袋类MHC基因型变异位于真实哺乳动物和鸟类之间,作为最小MHC编码,但组织与非哺乳动物的组织更接近,并且有袋动物的MHC I类基因在II类区域内扩增,产生独特的I / II类区域[11]
III类功能与I类和II类有很大不同,但其位点发生在人类的6号染色体上的其他两个类之间,并且经常在一起讨论。
| Class | Encoding | Expression |
|---|---|---|
| I | (1)肽结合蛋白,其选择用于抗原呈递的短氨基酸序列,以及(2)帮助抗原加工的分子(例如TAP和tapasin)。 | 一条名为α的链,其配体是CD8受体,特别是细胞毒性T细胞和NK细胞携带的抑制性受体。 |
| II | (1)肽结合蛋白和(2)协助抗原加载到MHC II类肽结合蛋白(例如MHC II DM,MHC II DQ,MHC II DR和MHC II DP)上的蛋白质。 | 两条链,称为α和β,其配体是辅助T细胞携带的CD4受体。 |
| III | 其他免疫蛋白,在抗原加工和呈递之外,例如补体级联的组分(例如,C2,C4,因子B),免疫信号传导的细胞因子(例如,TNF-α),以及从应激缓冲细胞的热休克蛋白 | 多种 |
1.3 人类 MHC 的遗传特点
HLA 基因复合体有多基因性、多态性、连锁不平衡和单元型遗传的特点,其 中最为突出的是多态性。
1.3.1 HLA 的多态性
多态性(polymorphism) 指在一个随机婚配的群体中单个基因座位上存在 两个以上的不同等位基因的现象。HLAI类和II类等位基因产物的表达具有共显 性特点,即同一个体中,一个基因座位上来自同源染色体的两个等位基因皆能得 到表达,因而一个个体通常拥有的经典I类和II类 HLA 等位基因产物有 12 种。
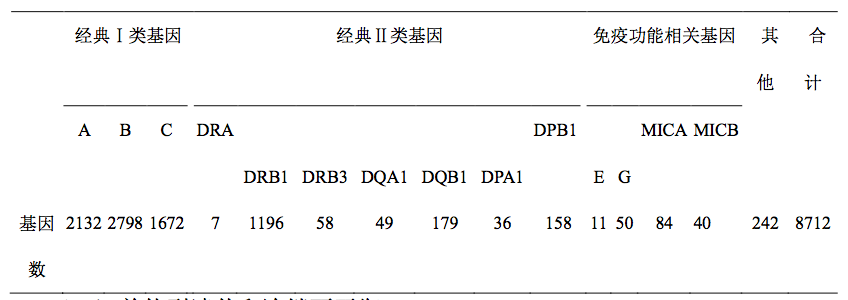
HLA 复合体是人体多态性最丰富的基因系统。截至 2012 年 10 月,已确定 的 HLA 等位基因总数达到 8712 个,其中等位基因数量最多的座位是 HLA-B (2798 个)。这表明,非亲缘关系个体间进行组织和器官移植,得到两个相同等 位基因的机会定然很低,因而,移植物会受到免疫排斥。表 6-1 提供了 HLA 主 要座位的等位基因数。
呈现多态性的 HLA 基因座位及已获正式命名的等位基因数(2012 年 10 月)
MHC等位基因以共显的方式表达。[13] 这意味着从父母双方遗传的等位基因(变体)同样表达:
- 每个人携带3个I类基因(HLA-A,HLA-B和HLA-C)中每一个的2个等位基因,因此可以表达六种不同类型的MHC-I(见图)。
- 在II类基因座中,每个人遗传一对HLA-DP基因(DPA1和DPB1,编码α和β链),一对基因HLA-DQ(DQA1和DQB1,用于α和β链),一个 基因HLA-DRα(DRA1)和一种或多种基因HLA-DRβ(DRB1和DRB3,-4或-5)。 这意味着一个杂合个体可以遗传六个或八个功能性II类等位基因,每个亲本三个或更多。 DQA2或DQB2的作用未经验证。 DRB2,DRB6,DRB7,DRB8和DRB9是假基因。
每个染色体中存在的一组等位基因称为MHC单倍型。在人类中,每个HLA等位基因都以数字命名。例如,对于给定的个体,他的单倍型可能是HLA-A2,HLA-B5,HLA-DR3等……每个杂合个体将具有两个MHC单倍型,每个来自父本和母本染色体。
MHC基因具有高度多态性;在群体内的不同个体中存在许多不同的等位基因。多态性如此之高,在混合群体中(非偶然性),除了同卵双胞胎外,没有两个个体具有完全相同的MHC分子组。
每个等位基因中的多态性区域位于肽接触的区域中。在MHC可以显示的所有肽中,只有一个子集与任何给定的HLA等位基因结合得足够强,因此通过携带每个基因的两个等位基因,可以呈现更大的肽组。[为什么?]
另一方面,在群体内,许多不同等位基因的存在确保总是存在具有特定MHC分子的个体,其能够加载正确的肽以识别特定的微生物。 MHC多态性的进化确保了群体不会屈服于新的病原体或突变的病原体,因为至少一些个体将能够产生足够的免疫应答来战胜病原体。 MHC分子(负责多态性)的变化是不同MHC分子遗传的结果,并且它们不是通过重组诱导的,因为抗原受体就是这种情况。
由于其基因中存在高水平的等位基因多样性,MHC也引起了许多进化生物学家的注意
1.3.2 单体型遗传
MHC 的单体型(haplotype)指同一染色体上紧密连锁的 MHC 等位基因的 组合。MHC 等位基因的构成和分布还有两个特点:1)等位基因的非随机性表达 和2)连锁不平衡
群体中各等位基因其实并不以相同的频率出 现。如 HLA-DRB1 和 HLA-DQB1 座位的等位基因数分别是 1196 和 179(见表 6-1),其中两个等位基因 DRB10901 和 DQB10701 在群体中的频率,按随机分 配的原则,应该是 0.1% (1/1196)和 0.6%(1/158),然而,在我国北方汉族人 群中它们的频率分别高达 15.6%和 21.9%。在斯堪的纳维亚白种人中,DRB1 和DQB1 基因座位中高频率分布的等位基因是 DRB10501 和 DQB10201。说明不 同人种中优势表达的等位基因及其组成的单体型可以不同。
1.3.3 连锁不平衡
不仅等位基因出现的频率不均一,两个等位基因同时出现 在一条染色体上的机会,往往也不是随机的。连锁不平衡(linkage disequilibrium) 指分属两个或两个以上基因座位的等位基因同时出现在一条染色体上的几率,高 于随机出现的频率。例如上面提到北方汉族人中高频率表达的等位基因 DRB10901 和 DQB10701 同时出现在一条染色体上的几率,按随机分配规律, 应是其频率的乘积为 3.4% (0.156×0.219=0.034),然而实际两者同时出现的频 率是 11.3%,为理论值的 3.3 倍。
非随机表达的等位基因和构成连锁不平衡的等位基因组成,因人种和地理族 群的不同而出现差异,属长期自然选择的结果。其意义在于,第一,可作为人种 种群基因结构的一个特征,追溯和分析人种的迁移和进化规律;第二,高频率表 达的等位基因如果与种群抵抗特定疾病相关,可以此开展疾病的诊断和防治;第 三,有利于寻找 HLA 相匹配的移植物供者。
- 连锁不平衡是一种遗传现象,其中两个等位基因以高于正常预期的频率一起被发现。
- 它是不同位点等位基因之间的非随机关联。例如,如果16%的人群具有特定的HLA-A抗原(A1), 10%的人群具有特定的HLA-B抗原(B8),则在同一染色体上发现A1与B8基因相关的机会由其基因频率的乘积给出(16% x 10% = 1.6%)。
- 在实践中,这种情况并不总是发生。如果A和B特异性的某些组合是随机的,那么它们发生的频率比预期的要高。A1和B8的组合在人群中出现的频率为8.8%,而预期的频率为1.6%,这种成对的特异性被称为连锁不平衡。这表明,处于连锁不平衡状态中的等位基因往往经常地连在一起,由此引入单体型的概念。单体型(haplotype)指的是染色体上MHC不同座位等位基因的特定组合。这样,某些单体型在群体中有可能呈现较高的频率,并较之单一座位的HLA基因型更能显示人种和地理位置特有的群体基因结构。同时,检测单体型比分析单一的等位基因频率,更有助于从无血缘关系人群中搜寻HLA相匹配的器官移植供者。
- 在高加索人中,HLA-A1、B8、DR3 (DRB10301)、DQ2 (DQB10201)单倍型在人群中高度保守。
- 在HLA II类中,这种现象非常明显,因此在检测前,可以使用特异性HLA-DR等位基因来预测HLA-DQ等位基因,准确度很高。6号染色体上的HLA等位基因顺序为DP-DQ-DR-B-Cw-A。
- 那些在实际中彼此最接近的等位基因通常具有最高的连锁不平衡。有可能某些单倍型在某些免疫学意义上是有利的,因此它们具有积极的选择优势。
二、HLA 分子的分布
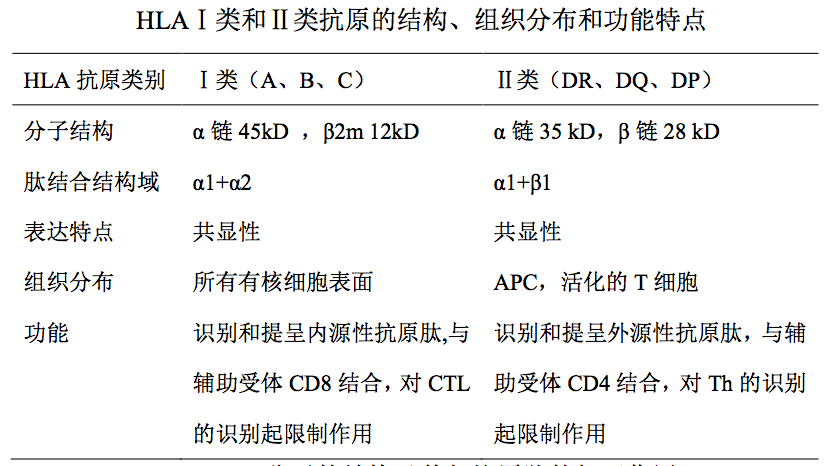
I类分子由重链(α 链)和 β2m 组成,分布于所有有核细胞表面。
II类分子由 α 链和 β 链组成,仅表达于淋巴组织中一些特定的细胞表面, 如专职性抗原提呈细胞(包括 B 细胞、巨噬细胞、树突状细胞)、胸腺上皮细 胞和活化的 T 细胞等。
三、HLA 分子的结构及其与抗原肽的相互作用
3.1 HLA 分子的结构
MHC蛋白具有免疫球蛋白样结构。
Class I
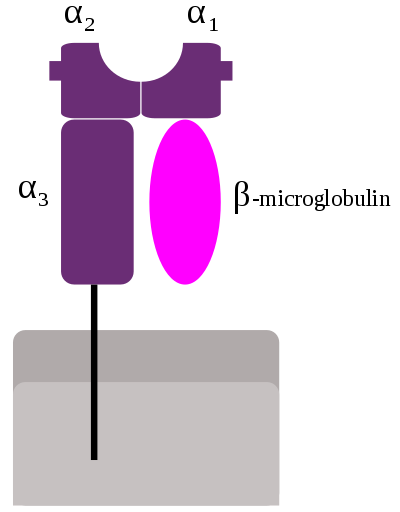
MHC I作为由三个结构域-α1,α2和α3组成的α链出现。 α1依赖于非MHC分子β2微球蛋白(在人15号染色体上编码)的单位。 α3结构域是跨膜的,将MHC I类分子锚定在细胞膜上。呈递的肽由肽结合沟的底部保持,位于α1/α2异二聚体的中心区域(由两个不相同的亚基组成的分子)。遗传编码和表达的氨基酸序列,即肽结合沟底板的残基序列决定了它所结合的特定肽残基。[12]
HLAI类分子是由重链(α 链)和轻链(β2m)以非共价键连接组成的异二 聚体糖蛋白。重链由胞外区、跨膜区和胞内区组成,胞外区有三个结构域 (α1、α2、α3)。I类分子的结构可以分为四个区:抗原肽结合区、免疫球蛋 白样区、跨膜区和胞质区。抗原肽结合区包括 α1 和 α2 两个结构域,二者构成 抗原肽结合槽,能与内源性抗原肽结合。免疫球蛋白样区主要包括重链 α3 结构 域和 β2m 微球蛋白:因二者氨基酸组成和免疫球蛋白恒定区具有高度同源性, 故称免疫球蛋白样区(Ig 样区),其中 α3 结构域是 CTL 表面 CD8 分子识别结 合的部位,β2m 微球蛋白与 α3 结构域结合有助于 HLAI类分子的表达和结合 稳定性。跨膜区含疏水性氨基酸残基,以 α 螺旋跨越脂质双层膜,并借此将 HLAI类分子锚定在细胞膜上。胞质区包括 α 链羧基末端约 30 个氨基酸残基, 内含可形成磷酸化的氨基酸序列,可能与细胞内外信号的传递相关。HLAI类 分子的抗原结合槽两端封闭,接纳的抗原肽长度有限,为 8~10 个氨基酸残 基。
Class II
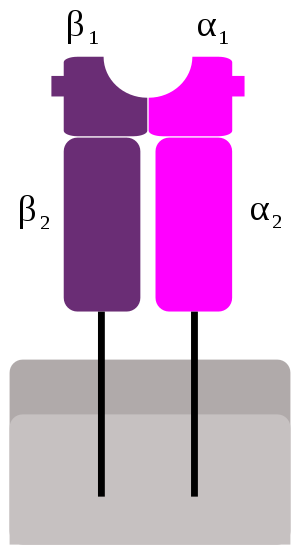
MHC II类由两条链α和β组成,每条链具有两个结构域-α1和α2以及β1和β2-各链分别具有跨膜结构域α2和β2,将MHC II类分子锚定在细胞膜上。[13] 肽结合沟由α1和β1的异二聚体形成。
人类中的MHC II类分子具有五至六个同种型。 经典分子将肽呈递给CD4 +淋巴细胞。 具有细胞内功能的非经典分子,附件不暴露在细胞膜上,而是暴露在溶酶体的内膜中,通常将抗原肽加载到经典的MHC II类分子上。
MHC II类蛋白质分子 经典MHC分子将表位呈递给CD8 + T淋巴细胞的TCR。非经典分子(MHC IB类)表现出有限的多态性,表达模式和呈递的抗原;该组被细分为在MHC基因座内编码的组(例如,HLA-E,-F,-G),以及那些不是(例如,应激配体如ULBPs,Rae1和H60);许多这些分子的抗原/配体仍然未知,但它们可以与CD8 + T细胞,NKT细胞和NK细胞中的每一种相互作用。
HLAII类分子是由 α 链(34KD)和 β 链(29KD)以非共价键结合组成的 异二聚体糖蛋白。α 链和 β 链为跨膜蛋白,均由胞外区、跨膜区和胞内区三部 分组成。II类分子的结构也可以分为四个区:抗原肽结合区、免疫球蛋白样 区、跨膜区和胞质区。抗原肽结合区包括 α1 和 β1 两个结构域,二者共同组成 抗原肽结合槽,能与外源性抗原肽结合。免疫球蛋白样区由 α2 和 β2 结构域组 成,其氨基酸组成和序列与免疫球蛋白恒定区具有高度同源性,故称免疫球蛋 白样区(Ig 样区),其中 β2 结构域是 Th 表面 CD4 分子识别结合的部位。跨膜 区和胞质区的氨基酸组成和功能与I类分子的类似,但胞质区内氨基酸残基数 明显少于I类分子。II类分子的抗原结合槽两端开放,进入槽内的抗原肽长度 变化较大,为 13~17 个氨基酸残基,甚至更长。
Class III
III类分子具有与I类和II类不同的生理作用,但在人类6号染色体的短臂中编码。III类分子包括具有免疫功能的几种分泌蛋白:补体系统的组分(如C2,C4和 B因子),细胞因子(如TNF-α,LTA和LTB)和热休克蛋白。
3.2 HLA 与抗原肽的相互作用
HLA 分子结合并提呈抗原肽供 TCR 识别。HLAI、II类分子接纳抗原肽 的结构,是位于该分子远膜端的抗原肽结合槽。HLA 的抗原肽结合槽与抗原肽 互补结合,其中有两个或两个以上与抗原肽结合的关键部位,称锚定位 (anchor position)。在该位置抗原肽与 HLA 分子结合的氨基酸残基组成称为 锚定残基(anchor residue)。
特定的 HLA 分子选择性地结合具有某共用基序的抗原肽,因此,结合显示 一定的专一性。这种相对专一性的意义在于:
- 具有某类 HLA 等位基因的个 体,可能由于 HLA 分子抗原肽结合槽与锚定残基不匹配,而对某种抗原不发生 或仅发生低水平免疫应答。
- 不同的 HLA 分子有可能提呈同一抗原的不同表 位,造成不同个体对同一抗原应答强度的差异。这实际上是 HLA 以其多态性参 与和调控免疫应答的一种重要机制。
- 实际上 HLA 分子对抗原肽的识别并非严 格的一对一关系,而是显示一定的包容性,造成一种类型的 HLA 分子可以识别 一群带有特定共用基序的肽段。这意味着,能够被某一 HLA 分子识别和提呈的 抗原肽,也可被该家族其他分子所提呈。这一点,对应用肽疫苗或 T 细胞疫苗 进行免疫预防和免疫治疗提供了便利。
四、HLA 分子的功能
在确定的三个MHC类别中,注意力通常集中在I类和II类。 通过与辅助T细胞表面上的CD4分子相互作用,MHC II类介导特异性免疫(也称为获得性免疫或适应性免疫)的建立。 通过与细胞毒性T细胞表面上的CD8分子相互作用,MHC I类介导受感染或恶性宿主细胞的破坏,特异性免疫的方面称为细胞免疫。 (特异性免疫的另一组是体液免疫,其与MHC的关系更为间接。)

MHC是组织抗原,其允许免疫系统(更具体地T细胞)结合,识别和耐受自身(自动识别)。 MHC也是细胞内肽的伴侣,其与MHC复合并呈递给T细胞受体(TCR)作为潜在的外来抗原。 在抗原结合亲和力和特异性以及信号转导有效性方面,MHC与TCR及其共受体相互作用以优化TCR-抗原相互作用的结合条件。
基本上,MHC-肽复合物是自身抗原/同种异体抗原的复合物。 结合后,T细胞原则上应该耐受自身抗原,但是当暴露于同种异体抗原时会激活。 当这个原则被破坏时就会出现疾病状态。
- 抗原呈递 ( Antigen presentation):MHC分子与T淋巴细胞上的T细胞受体和CD4 / CD8共同受体结合,并且MHC分子的肽结合沟中保持的抗原表位与TCR的可变Ig样结构域相互作用以触发 T细胞活化[9]
- 自身免疫反应(Autoimmune reaction):有一些MHC分子比其他人更容易增加自身免疫性疾病的风险。 HLA-B27就是一个例子。 尚不清楚HLA-B27组织类型究竟是如何增加强直性脊柱炎和其他相关炎性疾病的风险,但已经假设了涉及异常抗原呈递或T细胞活化的机制。
- 组织同种异体识别 (Tissue allorecognition):与肽表位复合的MHC分子基本上是TCR的配体。 T细胞通过与任何MHC分子的肽结合沟结合而被激活,在胸腺阳性选择期间T细胞未被训练识别。
淋巴细胞 (Lymphocytes)
作为白细胞的谱系,淋巴细胞存在于外周淋巴组织中,包括淋巴滤泡和淋巴结,并且包括B细胞,T细胞和天然杀伤细胞(NK细胞)。具体起作用的B细胞分泌抗体分子,但不结合MHC。特异性起作用的T细胞以及天然起作用的NK细胞与MHC相互作用。 NK细胞表达与MHC I分子结合的杀伤Ig样受体(KIR),并通过ITIM(免疫受体酪氨酸抑制基序)募集和激活蛋白酪氨酸磷酸酶发出信号。这意味着与激活Tc淋巴细胞的CD8 / TCR相互作用相反,NK细胞在与MHC I结合时变得失活。当MHC I类表达低时,通常在病毒感染或肿瘤发生期间异常细胞功能的情况下,NK细胞丢失抑制性KIR信号并触发异常细胞的程序性细胞死亡。因此,NK细胞通过促进肿瘤监测来帮助预防癌细胞的进展
MHC II Class
MHC II类可以由所有细胞类型有条件地表达,但通常仅在专职抗原呈递细胞(APC)上发生:巨噬细胞,B细胞,尤其是树突细胞(DC)。 APC吸收抗原蛋白,进行抗原加工,并返回其分子部分 - 称为表位的部分 - 并将其显示在偶联于MHC II类分子(抗原呈递)的APC表面上。在细胞表面,表位可被免疫结构如T细胞受体(TCR)识别。与表位结合的分子区域是互补位(paratope)。
在辅助T细胞的表面上是CD4受体,以及TCR。当幼稚辅助T细胞的CD4分子与APC的MHC II类分子对接时,其TCR可以与MHC II类内偶联的表位相遇并被其印记。这个事件引发了幼稚辅助T细胞。根据当地的环境,即APC在微环境中分泌的细胞因子的平衡,幼稚辅助T细胞(Th0)极化为记忆Th细胞或表型的效应Th细胞1型(Th1),类型如图2所示,Th(Th2),17型(Th17)或调节/抑制子(Treg)是Th细胞的终末分化。
因此,如果APC主要向Treg细胞极化Th0细胞,则MHC II类介导免疫 - 抗原的免疫耐受性。初次暴露于抗原期间的极化是确定许多慢性疾病(例如炎性肠病和哮喘)的关键,通过使得记忆Th细胞在二次暴露于类似抗原时触发记忆回忆时的免疫应答发生偏斜。 (B细胞表达MHC II类以将抗原呈递给Th0,但是当它们的B细胞受体结合匹配的表位,不是由MHC介导的相互作用时,这些活化的B细胞分泌可溶性免疫球蛋白:介导体液免疫的抗体分子。)
MHC I Class
MHC I类发生在所有有核细胞上,也发生在血小板中 - 实质上是所有细胞,但是红细胞。 它将表位呈递给杀伤性T细胞,也称为细胞毒性T淋巴细胞(CTL)。 除TCR外,CTL还表达CD8受体。 当CTL的CD8受体与MHC I类分子对接时,如果CTL的TCR与MHC I类分子内的表位相符,则CTL通过细胞凋亡触发细胞经历程序性细胞死亡。 因此,MHC I类有助于介导细胞免疫,这是解决细胞内病原体的主要手段,例如病毒和一些细菌,包括细菌L形式,细菌属Mycoplasma和细菌属立克次氏体。 在人类中,MHC I类包含HLA-A,HLA-B和HLA-C分子。
4.1 作为抗原提呈分子参与适应性免疫应答
经典的 MHCI类和II类分子通过提呈抗原肽而激活 T 淋巴细胞,参与适应 性免疫应答。这是 MHC 主要的生物学功能。由此派生出特异性免疫应答中与这 一功能相关的一系列表现。
- MHC 限制性(MHC restriction) 指 T 细胞以其 TCR 对抗原肽和自身 MHC 分子进行双重识别,即 T 细胞只能识别自身 MHC 分子提呈的抗原肽,其 中 CD4+Th 细胞识别II类分子提呈的外源性抗原肽,CD8+CTL 识别I类分子提 呈的内源性抗原肽。
- 参与 T 细胞在胸腺中的选择和分化。
- 决定疾病易感性的个体差异 近代确认的疾病关联原发成分,许多属于 特定的 HLA 等位基因(或与之紧密连锁的疾病易感基因),以及这些基因的产 物。这些基因的作用和 HLA 分子的抗原提呈功能密切相关。
- 参与构成种群基因结构的异质性 由于不同 MHC 分子加工提呈的抗原 肽往往不同,这一特点赋予不同个体抗病能力的差异,这在群体水平有助于增强 物种的适应能力。
抗原处理和呈现 Antigen processing and presentation
肽由两种经典途径加工和呈现:
在MHC II类中,吞噬细胞如巨噬细胞和未成熟的树突细胞通过吞噬作用吸收实体进入吞噬体 - 尽管B细胞表现出更广泛的胞吞内吞作用 - 其与溶酶体融合,其酸性酶将吸收的蛋白质切割成许多不同的肽。通过与宿主承载的特定MHC II类变体的分子相互作用中的物理化学动力学,在宿主的基因组中编码,特定肽表现出免疫优势并加载到MHC II类分子上。这些被贩运到细胞表面并在细胞表面外化。[14]
在MHC I类中,任何有核细胞通常呈现胞质肽,主要是源自蛋白质周转和核糖体缺陷产物的自身肽。在病毒感染,细胞内微生物感染或癌转化过程中,蛋白质体中降解的这些蛋白质也被加载到MHC I类分子上并显示在细胞表面上。 T淋巴细胞可以检测到显示在MHC分子的0.1%-1%的肽。
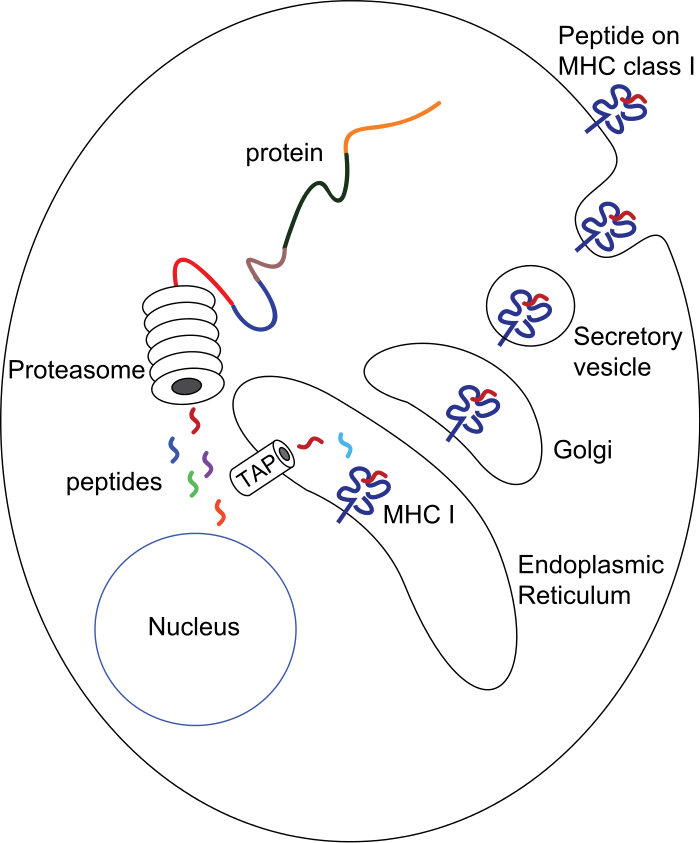
MHC class I pathway: Proteins in the cytosol are degraded by the proteasome, liberating peptides internalized by TAP channel in the endoplasmic reticulum, there associating with MHC-I molecules freshly synthesized. MHC-I/peptide complexes enter Golgi apparatus, are glycosylated, enter secretory vesicles, fuse with the cell membrane, and externalize on the cell membrane interacting with T lymphocytes.
表2.抗原加工途径的特征
| Characteristic | MHC-I pathway | MHC-II pathway |
|---|---|---|
| 稳定的肽-MHC复合物的组成 | 多态链α和β2微球蛋白,肽与α链结合 | 多态链α和β,肽与两者结合 |
| 抗原呈递细胞的类型(APC) | 所有有核细胞 | 树突细胞,单核吞噬细胞,B淋巴细胞,一些内皮细胞,胸腺上皮细胞 |
| T淋巴细胞能够反应 | 细胞毒性T淋巴细胞(CD8 +) | 辅助T淋巴细胞(CD4 +) |
| 抗原蛋白的起源 | 细胞溶质蛋白(主要由细胞合成;也可能通过吞噬体从细胞外介质进入) | 存在于内体或溶酶体中的蛋白质(大多数是从细胞外培养基中内化的) |
| 负责肽生成的酶 | 细胞溶质蛋白酶体 | 来自内体和溶酶体的蛋白酶(例如,组织蛋白酶) |
| 将肽加载到MHC分子上的位置 | 内质网 | 专门的囊泡隔室 |
| 涉及运输肽并将它们加载到MHC分子上的分子 | TAP(与抗原加工相关的转运蛋白) | DM,不变链 |
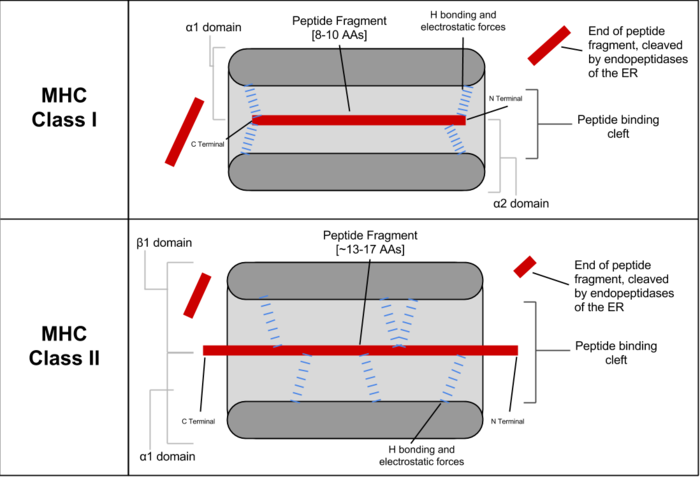
Peptide binding for Class I and Class II MHC molecules, showing the binding of peptides between the alpha-helix walls, upon a beta-sheet base. The difference in binding positions is shown. Class I primarily makes contact with backbone residues at the Carboxy and amino terminal regions, while Class II primarily makes contacts along the length of the residue backbone. The precise location of binding residues is determined by the MHC allele.[15]
4.2 作为调节分子参与固有免疫应答
HLA 中的免疫功能相关基因参与对固有免疫应答的调控主要表现在以下方 面。
- 经典的III类基因 为补体成分编码,参与炎症反应和对病原体的杀伤,与 免疫性疾病的发生有关。
- 非经典I类基因和 MICA 基因 产物可作为配体分子,以不同的亲和力 结合激活性和抑制性受体,调节 NK 细胞和部分杀伤细胞的活性。
- 炎症相关基因 参与启动和调控炎症反应,并在应激反应中发挥作用。
五、HLA 与临床
5.1 HLA 与器官移植
同种异体器官移植物存活率的高低主要取决于供体和受体之间的 HLA 型别 相合的程度。同卵双生个体(HLA 完全相同)间进行器官和骨髓移植不发生移 植排斥反应,移植物可长期存活。同胞间出现 HLA 基因完全相同的概率为 25%, 器官移植时应首先从兄弟姐妹中寻找相同配型。在一个单倍型相同的同胞或父母 与子女间进行器官移植,其存活率高于无亲缘关系供受体间器官移植的存活率。 因此,通常器官移植物存活率由高到低的顺序是:同卵双生>同胞>亲属>无亲 缘关系。在肾移植中,HLA 各位点基因配型的重要性依次为 HLA-DR>HLA-B >HLA-A;在骨髓移植中,只有在供体和受体 HLA 单体型完全相同情况下才容 易获得成功。临床上测定血清中可溶性 HLA 分子的含量,有助于监测移植物的 排斥危象。
5.2 HLA 与输血反应
临床上多次接受输血的病人会发生非溶血性输血反应。患者主要表现为发热、 白细胞减少和荨麻疹等临床症状。此类输血反应主要与患者血液中产生针对供体 HLAI类抗原特异性抗体有关。患者在多次输血过程中若再次接受同一供体血液, 其体内相应 HLA 特异性抗体即能与供体白细胞/血小板表面的 HLAI类分子结 合,进而激活补体使上述细胞溶解破坏发生非溶血性输血反应。因此,对多次接 受输血者应注意避免反复选择同一供血者的血液。
5.3 HLA 分子的异常表达和临床疾病
所有有核细胞表面表达 HLAI类分子,但肿瘤细胞I类分子的表达往往减 弱甚至缺如,以致不能有效地激活特异性 CD8+CTL,造成肿瘤逃脱免疫监视。 与此相反,某些自身免疫病中,原先不表达 HLAII类分子的上皮细胞,可被诱 导表达II类分子,如胰岛素依赖性糖尿病中的胰岛 β 细胞、乳糜泻中的肠道细 胞、萎缩性胃炎中的胃壁细胞等。上述异常表达的机制及其免疫病理学意义未明, 可能和它们促进免疫细胞的过度活化有关。
5.4 HLA 和疾病关联
(一)HLA 是人体对疾病易感的主要免疫遗传学成分
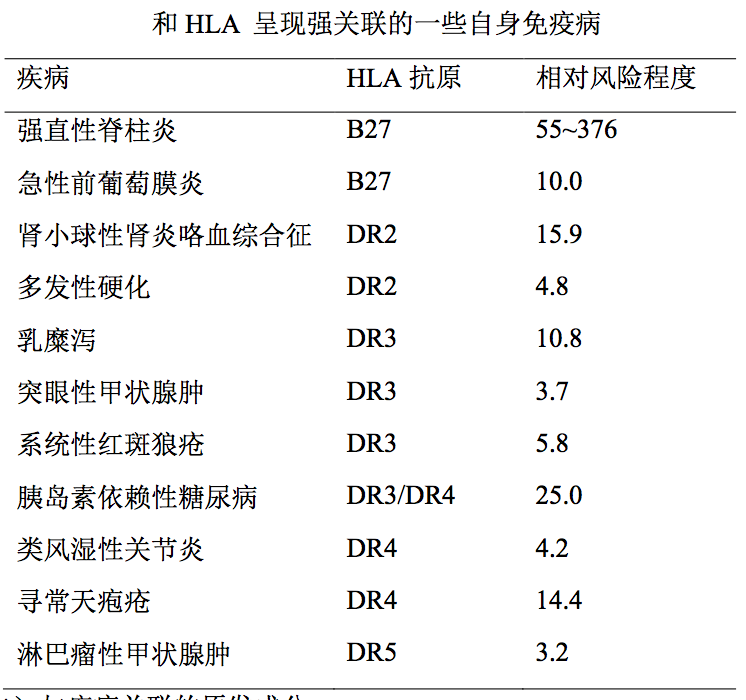
带有某些特定 HLA 型别的个体易患某一疾病(称为阳性关联)或对该疾病 有较强的抵抗力(称为阴性关联)皆纳入 HLA 和疾病关联的范畴。这一关联, 是通过对患病人群和健康人群作 HLA 分型后,用统计学方法加以判别的。典型 例子是强直性脊柱炎(AS)。发现患者中 HLA-B27 抗原阳性率高达 58%~97%, 而在健康对照人群中仅为 1%~8%,由此确定强直性脊椎炎和 HLA-B27 属阳性 关联。换言之,带有 B27 等位基因的个体易于患强直性脊椎炎。经计算,其相对 风险程度(RR)为 55~376(因不同人种而异),即 B27 阳性个体较之 B27 阴性 个体罹患 AS 的机会要大 55 倍到 376 倍。表明 HLA-B27 是决定强直性脊柱炎疾 病易感性的关键遗传因素。迄今,记录在案和 HLA 关联的疾病达 500 余种,大 部分为自身免疫病。
(二)与疾病关联的原发成分
B27 和 AS 的关联是一个十分典型的例子,已确定 B27 是原发关联成分。对 其它疾病并非如此,因而 HLA 和疾病关联的研究一般包括两个方面:一是寻找 关联的原发成分,包括从分子水平确定起核心作用的 HLA 等位基因,排除因连 锁不平衡而出现的次级关联,并确定相应抗原的关键性表位;二是阐明关联机制。 下面举例说明一些已获进展的疾病。
- 类风湿性关节炎(RA) 这是一种以DR4为主要关联成分的自身免疫性 疾病。DR4 等位基因分子 67~74 位氨基酸残基序列为 LLEQRRAA,该等位基 因即属易感成分。
- 乳糜泻(CD) 是由面粉麸质引起的小肠黏膜吸收不良综合征。已阐明 该病的原发关联成分是和 DR3 呈现连锁不平衡的 HLA-DQA10501 及 DQB10201。这两个等位基因可以分别采用顺式互补(在同一条染色体上)或反 式互补(在两条同源染色体上)两种形式,决定易感性。
- 胰岛素依赖性糖尿病(IDDM) 原发关联成分并非表 6-3 所列的 DR 基 因,而是与之处于连锁不平衡的 DQ 基因即 DQA10301 和 DQB10201。两个 DQ 等位基因也呈现顺、反两种互补形式。另外存在一些易感程度较低的单元型 组合以及抵抗性等位基因。
- 多发性硬化症(MS) 一种神经系统脱髓鞘性疾病,相关联的成分已确 定为 DR2 中的 DRB11501,并有 DRB30101 参与。
5.5 HLA 与法医学和亲子鉴定
HLA 系统所显示的多基因性和多态性,意味着两个无亲缘关系个体之间, 在所有 HLA 基因座位上拥有相同等位基因的机会几乎等于零。而且,每个人所 拥有的 HLA 等位基因型别一般终身不变。这意味着特定等位基因及其以共显性 形式表达的产物,可以成为不同个体用以显示个体性(individuality)的遗传标志。 据此,HLA 基因分型已在法医学上被广泛地用于亲子鉴定和对死亡者“验明正 身”。
六、其他
在细胞中,宿主自身表型或其他生物实体的蛋白质分子不断合成和降解。 细胞表面的每个MHC分子都显示一种蛋白质的分子部分,称为表位(epitpe)[2]。 所呈递的抗原可以是自身的或非自身的,从而防止生物体的免疫系统靶向其自身细胞。 整体而言,MHC群体就像表盘,表明细胞内蛋白质的平衡。
MHC基因家族分为三个亚组:I类,II类和III类。 I类MHC分子具有β2亚基,其仅能被CD8共同受体识别。 II类MHC分子具有β1和β2亚基,并且可被CD4共受体识别。 以这种方式,MHC分子陪伴哪种类型的淋巴细胞可以高亲和力结合给定抗原,因为不同的淋巴细胞表达不同的T细胞受体(TCR)共同受体。
由MHC I类和II类介导的抗原呈递的多样性以至少三种方式实现:(1)生物体的MHC谱系是多基因的(通过多个相互作用的基因); (2)MHC表达是共显性的(来自两组遗传等位基因); (3)MHC基因变异是高度多态的(在物种内从生物体到生物体各不相同)。[3] 在雄性小鼠中观察到主要的组织相容性复合体和性别选择,使得具有不同MHC的雌性的配偶选择,从而证明性选择。[4] 此外,至少对于MHC I呈现,已有证据表明抗原肽剪接可以结合来自不同蛋白质的肽,大大增加抗原多样性
6.1 T淋巴细胞识别限制
在它们在胸腺中的发育中,选择T淋巴细胞来识别宿主的MHC分子,但不识别其他自身抗原。选择后,每个T淋巴细胞显示出双重特异性:TCR识别自身MHC,但仅识别非自身抗原。
通过称为阳性选择的过程,在胸腺中的淋巴细胞发育期间发生MHC限制。没有接受阳性存活信号的T细胞 - 主要由呈现与MHC分子结合的自身肽的胸腺上皮细胞 - 介导其TCR经历细胞凋亡。阳性选择确保成熟T细胞可以在功能上识别外周(即身体其他部位)的MHC分子。
T淋巴细胞的TCR仅识别仅有肽的连续表位,也称为线性表位,并且仅在MHC分子内偶联时才识别。 (然而,由活化的B细胞分泌的抗体分子连接不同的表位 - 肽,脂质,碳水化合物和核酸 - 并识别具有三维结构的构象表位。)
6.2 在性伴侣选择中
MHC分子能够对宿主细胞中蛋白质分子群体进行免疫系统监测,并且更大的MHC多样性允许更大的抗原呈递多样性。 1976年,Yamazaki等人证明了雄性小鼠对不同MHC雌性的性选择配偶选择。用鱼获得了类似的结果。[16]一些数据发现不同MHC基因的人类夫妇早期妊娠丢失率较低[17]。
MHC可能与某些人群中的配偶选择有关,这一理论得到了Ober及其同事1997年的研究[18]以及Chaix及其同事在2008年的研究。[19]然而,后者的研究结果一直存在争议。[20]如果存在,该现象可能是由嗅觉介导的,因为MHC表型似乎强烈地涉及来自汗液的化合物的感知气味的强度和舒适性。脂肪酸酯 - 如十一酸甲酯,癸酸甲酯,壬酸甲酯,辛酸甲酯和己酸甲酯 - 显示出与MHC的强烈联系。[21]
1995年,Claus Wedekind发现,在一群女大学生中,他们闻到男学生穿的T恤两晚(没有除臭剂,古龙水或香皂),到目前为止,大多数女性都选择了不同MHC男士穿的衬衫,如果妇女使用口服避孕药,则优先考虑。[22] 2002年实验的结果同样表明HLA相关的气味会影响气味偏好并可能调节社会线索。[23] 2005年,在一组58名受试者中,女性在接受像他们自己的MHC时更加犹豫不决,[24]尽管使用口服避孕药,但女性并没有特别偏好。[25]没有研究表明气味偏好决定配偶选择的程度(反之亦然)。
6.3 进化多样性
大多数哺乳动物具有与人类相似的MHC变体,其具有很大的等位基因多样性,特别是在九个经典基因中 - 似乎主要是由于基因重复 - 尽管人类MHC区域具有许多假基因。最多样化的基因座,即HLA-A,HLA-B和HLA-DRB1,分别具有大约1000,1600和870个已知等位基因[需要引证]。[26]许多HLA等位基因是古老的,有时与黑猩猩MHC等位基因的同源性高于同一基因的一些其他人类等位基因。
MHC等位基因多样性挑战了进化生物学家的解释。大多数平衡选择(参见多态性(生物学)),这是任何自然选择过程,其中没有单个等位基因绝对最适合,例如频率依赖性选择和杂合子优势。最近的模型表明,仅通过杂合子优势,大量等位基因难以置信。[引证需要]
致病共同进化是一种反假设,认为常见的等位基因处于最大的致病压力下,驱使正常选择不常见的等位基因 - 移动目标,即病原体。由于先前常见等位基因的致病压力降低,它们在群体中的频率稳定,并且仍然在大量群体中循环。尽管在群体水平上具有很好的MHC多态性,但个体最多携带18个MHC I或II等位基因。
在猎豹(Acinonyx jubatus),[27] Eurasian beaver(Castor fiber),[28]和大熊猫(Ailuropoda melanoleuca)中观察到相对较低的MHC多样性。[29] 2007年,低MHC多样性被认为是塔斯马尼亚恶魔(Sarcophilus harrisii)疾病易感性的一个角色,原产于孤立的塔斯马尼亚岛,因此参与恶魔面部肿瘤疾病的传染性肿瘤的抗原似乎被认为是自我抗原。[30]为了抵消近亲繁殖,已经提出了维持濒危物种和圈养动物种群遗传多样性的努力。
6.4 在移植排斥 In transplant rejection
在移植过程中,如器官或干细胞,MHC分子本身作为抗原并且可以在受体中引起免疫应答,从而引起移植排斥。 MHC分子在它们在不同菌株的小鼠之间的移植排斥中起作用后被鉴定和命名,尽管它花了20多年来阐明MHC在将肽抗原呈递给细胞毒性T淋巴细胞(CTL)中的作用。[31]
每个人类细胞表达6个MHC I类等位基因(来自每个亲本的一个HLA-A,-B和-C等位基因)和6到8个MHC II类等位基因(一个HLA-DP和-DQ,以及一个或两个HLA-DR)来自每个父母,以及这些的组合)。人群中的MHC变异高,HLA-A基因的至少350个等位基因,HLA-B的620个等位基因,DR的400个等位基因和DQ的90个等位基因。任何两个不是同卵双胞胎的人都会表达不同的MHC分子。所有MHC分子均可介导移植排斥,但HLA-C和HLA-DP显示低多态性,似乎最不重要。
当在胸腺中成熟时,选择T淋巴细胞的TCR无法识别自身抗原,但是T淋巴细胞可以对供体MHC的肽结合沟反应,MHC的可变区保持呈递的抗原表位以通过TCR识别,匹配互补位。受体的T淋巴细胞将不相容的肽结合沟作为非自身抗原。 T淋巴细胞识别外来MHC为自我是同种异体识别
移植排斥反应已知有多种类型由MHC(HLA)介导:
- 在移植前,受体已经通过先前的输血(包括表达HLA分子的淋巴细胞的供体组织),通过在怀孕期间产生的抗HLA(针对由父亲的HLA显示)进行抗HLA抗体,发生超急性排斥。胎儿),或通过以前的移植;
- 当受体的T淋巴细胞被供体组织激活时,发生急性细胞排斥,通过诸如CD8细胞的直接细胞毒性等机制引起损伤。
- 当受体的抗HLA抗体形成针对移植组织的内皮细胞上存在的HLA分子时,发生急性体液排斥和慢性功能障碍。
在所有上述情况中,免疫是针对移植器官,维持病变。潜在供体细胞和受体血清之间的交叉反应试验旨在检测识别供体HLA分子的潜在受体中预先形成的抗HLA抗体的存在,从而防止超急性排斥。在正常情况下,评估HLA-A,-B和-DR分子之间的相容性。不相容性越高,五年存活率越低。捐助者信息全球数据库加强了对兼容捐助者的搜索
七、其他问题
7.1 HLA命名法
- HLA命名法是从最初的血清学名称发展而来的,蛋白质的多态性最初是由抗体反应模式定义的,现代定义利用DNA序列来定义等位基因。目前的命名法是在1987年第十届国际组织相容性研讨会上推荐的,并在1990年进行了轻微修改。
- 每条染色体在每个个体中被发现两次(二倍体),因此一个个体的正常组织类型将涉及12个HLA抗原(来自每个亲本的3个HLA I类位点(A、B和C)和来自每个亲本的3个HLA II类位点(DR、DQ和DP)。
- HLA-DM和HLA-DO不是高度多态性的,也不是分型的,这12种抗原是共显性遗传的。
- 一个人的MHC表型描述了这个人携带的等位基因,而不涉及遗传。例如,有人可能会输入HLA-A1,-A3;B7,B8;Cw2,Cw4;DR15,DR4;DQ3,DQ6;DP4,DP4。
- 单倍型是从父母一方遗传的一组HLA抗原。例如,上述HLA类型的人的母亲可能有HLA- A3,-A69;B7,B45;Cw4,Cw9;DR15,DR17;DQ6,DQ2;DP2,DP4。因此,A3、B7、Cw4、DR15、DQ6和DP4都是从母亲遗传给上面的孩子的,这组抗原是单倍型。
- 尽管在每个表达位点上有大量的等位基因,但在人群中观察到的单倍型数量远小于理论预期。这是因为某些等位基因倾向于在同一单倍型上一起出现,而不是随机分离,这叫做连杆不平衡。

7.2 HLA命名规则
- HLA前缀位于所有抗原或等位基因之前。
- 大写字母表示特定的位点(A、B、C或D)。所有在D区域的基因都以字母D开头,然后用第二个字母表示D的子区域(DR、DQ、DP、DM或DO)。
- 接下来确定编码特定II类肽链的位点(A1, A2, B1和B2)。希腊字母用于蛋白质命名,而拉丁大写字母用于基因/等位基因命名,即DRp1与DRB1。
- 特异等位基因用“ * ”表示,第一个两位数字表示最密切相关的血清学特异性,第二个两位数字表示唯一的等位基因。例如,血清学定义的HLA-A2特异性实际上包括77种不同的变异等位基因,这些等位基因现在被称为HLA-A *02:01到 *02:77。
- 一些等位基因有第三个两位数字(HLA-B *35:01:01和HLA-B *35:01:02),这表明这两个变体的差异是由沉默的核苷酸取代而不是氨基酸序列。

HLA等位基因命名包括基因名称(A、B、DR等)、等位基因族、等位基因亚型、非编码区的多态性、内含子区的多态性等信息。对某一个等位基因,先写出座位名,下接*,再用4个数字代表这个等位基因的名字。例如HLA-A * 02:01,其中A指HLA基因座位, 后面的4位数中的前两位是指这个等位基因相应的血清学特异性,这个例子中血清学特异性就是A02;后两位数则代表该等位基因序号(或者叫等位基因亚型),如果两个等位基因的4个数字都不同,则表示这两个等位基因编码的蛋白不同。如果一个等位基因在不同个体间仅在非编码区不同表明出现无义突变,所编码的蛋白分子不变,则在等位基因后再加一位或两位数字以示区别。如HLA-A02:01:01与HLA-A*02:01:02代表了非编码区不同基因序列,但编码出来的氨基酸序列是完全相同的。其实还可以往后写,但基本上就没有必要去关注,所以分辨率的基本达到02:01这种程度就足够。
参考资料
- https://en.wikipedia.org/wiki/Major_histocompatibility_complex
- 《医学免疫学》课件 新乡医学院 宋向凤、张国俊、徐春阳、孙爱平、孙书明、赵铁锁等老师
- https://mp.weixin.qq.com/s/RKm4CtpLo9LMgh6hxhzpeQ
- Immunology IV: Clinical Applications in Health and Disease. I Care Press, Bethesda, MD, 2012.
- https://www.immunopaedia.org.za/immunology/basics/
- 《医学免疫学》人卫第9版
