【2.2.1.1】RIG-I样受体(RLRs,RIG⁃I、MDA5、LGP2 )
机体的免疫反应包括先天存在的天然免疫反应和高度特异的适应性免疫反应,二者相互作用共同抵御病原微生物的侵袭。在天然免疫反应中,宿主细胞的特异性受体能够识别病原相关分子模式(pathogen associated molecular patterns,PAMPs),从而诱导I型干扰素(IFNs)和炎性细胞因子产生。宿主细胞特异性受体被称为模式识别受体(pattern recognition receptors,PRRs),主要包括:
- Toll样受体(toll-like receptors,TLRs)
- RIG-I样受体(RIG-I like receptors,RLRs)
- NOD样受体(NOD-like receptors,NLRs)
- C型凝集素样受体(C-type lectin receptors,CLRs)
- 胞浆DNA受体(cytosolic DNA receptors,CDRs)
RLRs作为细胞质中重要的PRRs,能够识别双链RNA病毒,是抗病毒天然免疫与适应性免疫非常重要的组成部分。其家族有3个成员:
- 遗传与生理实验分子(laboratory of genetics and physiology 2,LGP2)
- 视黄酸诱导基因I(retinoic acid inducible gene I,RIG-I)
- 黑色素瘤分化相关基因 5(melanoma differentiation associated gene 5,MDA5)。
近年来,对RIG-I、MDA5的功能研究很透彻,RIG-I、MDA5可分别识别短链、长链RNA,从而诱导下游IFNs产生;而LGP2的功能尚不清楚,因此进一步探究LGP2的确切功能很有必要。
一、RIG⁃I、MDA5介导的信号通路
RLRs家庭成员(RIG-I、MDA5和LGP2),在分子结构上具有同源性,都具有识别病毒dsRNA与水解腺嘌呤核苷三磷酸(ATP)的能力,但在酶活性、内在识别RNA能力和活化机制等方面存在差异。在正常细胞中,RLRs表达量较低,但在被病毒感染的细胞中,可被IFNs激活。
- RIG-I作为胞质中识别病毒dsRNA的传感元件,能够优先识别短链dsRNA(<300 bp)和5’端含有三磷酸基团的dsRNA(5'-triphosphorylated RNA,5'-ppp-dsRNA)。
- MDA5能够优先识别长链dsRNA(>1 000 bp),也能识别人工合成的dsRNA类似物聚肌胞苷酸(polycytidylic acid,polyI:C)。
RIG-I、MDA5活化后都可诱导IFNs表达,从而激活抗病毒免疫反应。
RIG-I、MDA5具有3个结构域:
- N端串联Caspase活化与聚合区域(caspase activation and recruitment domains,CARDs)、
- 中间RNA解旋酶结构域(RNA helicase domain)
- C端调控或抑制结构域(C-terminal domain,CTD或repressor domain,RD)。
宿主细胞处于静息状态时,RIG-I、MDA5的CARDs区与自身RD结构域结合,此时二者处于非活化状态。病毒感染宿主细胞时,RIG-I、MDA5的CTD结构域识别病毒dsRNA并且发生构象上的改变,致使二者CARDs区都被激活,进而活化下游含CARDs区的信号接头蛋白——线粒体抗病毒信号蛋白(mitochondrial antiviral signaling,MAVS或IPS-1/VISA/Cardiff)。活化的 MAVS 能够激活TRAF3(TNF receptor associated factor 3)和TRAF2/6(TNF receptor associated factor 2/6)。其中:TRAF3 进一步激活TBK1、IKKε蛋白激酶,二者通过磷酸化激活转录因子IRF3/7(interferon regulatory Factor 3/7)诱导IFNs产生;TRAF2/6通过与RIP1(receptor interacting protein 1)、TRADD(TNF receptor type 1 associated death domain protein)、Caspase 8/10相互作用进一步激活IKK复合物(IKKα/IKKβ/IKKγ),从而激活核因子κB蛋白(nuclear factor kappa-B,NF-κB),诱导产生炎性细胞因子。IFNs、炎性细胞因子和其他抗病毒产物共同作用产生抗病毒效应。图1是依据文献绘制的LGP2双向调控RLRs介导的天然免疫信号通路。
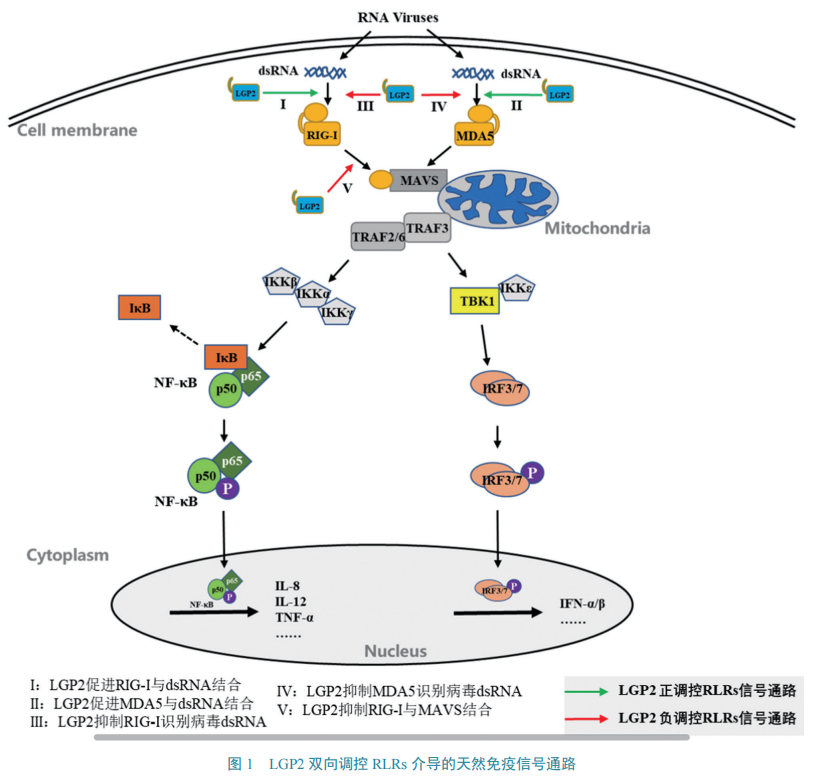
二、LGP2 结构特征及其在RLRs介导信号通路中的作用
根据遗传学与生理学研究,人类LGP2蛋白由17号染色体上的DHX58基因编码。DHX58基因全长共11 329 bp,包含14个外显子与13个内含子。LGP2蛋白由678个氨基酸组成,相对分子质量约为75 kDa。LGP2作为RLRs家族的一员,与RIG-I、MDA5具有显著的结构相似性,但其对病毒dsRNA的识别能力与RIG-I、MDA5有所差异。
LGP2具有与RIG-I、MDA5相同的DExD/H盒、中间解旋酶结构域和CTD结构域,这使其可以识别并结合病毒dsRNA。但与RIG-I、MDA5相比,LGP2 缺少CARDs区,因此不能与下游信号分子结合。但LGP2可通过强大的结合病毒dsRNA能力来调控RIG-I、MDA5介导的信号通路。早期细胞转染试验研究表明,LGP2能负调控RIG-I、MDA5介导的信号通路。此后利用LGP2缺陷(LGP2-/-)小鼠研究发现,LGP2的ATP酶结构域可以促进RIG-I、MDA5对病毒dsRNA的识别,从而正向调控RIG-I、MDA5介导的信号通路(图1:I)。进一步研究发现:LGP2不仅具有双向调控作用而且具有一定的浓度依赖性,当细胞处于感染早期时,低浓度水平的LGP2可以促进MDA5活化来抵抗病毒感染;而在感染后期,当LGP2浓度升高时,则会抑制RIG-I、MDA5的活化。
近年来,生理性低浓度的LGP2对MDA5的正向调控作用已被广泛证实,其作用的生化机制与分子机制逐渐清晰。在RLRs家族成员中,MDA5可与长链dsRNA缓慢形成不稳定的纤丝,因此MDA5与病毒dsRNA结合能力较弱;而LGP2结合dsRNA与5'-PPP和RNA的长度无关,且不形成纤丝,因此LGP2与病毒dsRNA结合能力最强。另外,LGP2的ATP水解酶活性区域不仅可以增强MDA5与dsRNA 的初始结合,还可以使MDA5在dsRNA上形成短纤丝并保持稳定,从而提高MDA5的生理活性(图1:II)。Li等的研究同样发现,猪LGP2可与MDA5相互作用,增强MDA5与dsRNA的结合能力,从而正向调控MDA5介导的信号通路。
另一方面,LGP2能够通过多种方式抑制RIG-I介导的信号通路(图1:III)。其C端RD结构域可与RIG-I的RD结构域结合形成异二聚体复合物,抑制RIG-I多聚化,从而抑制RIG-I识别病毒dsRNA;LGP2也可抑制RIG-I与MAVS结合(图1:V),进而抑制RIG-I的信号传导。另外,LGP2 还可通过抑制RIG-I下游转录因子IRF3的活化从而发挥潜在的负调控。近年来,有研究报道称,多种RNA解旋酶,例如DDX3、DHX29、DHX36和DDX60,都参与了对RIG-I、MDA5的正向调控作用。猪LGP2产生的效应与上述RNA解旋酶类似,很可能也正向调控RIG-I、MDA5介导的信号通路。
在不同物种中,LGP2对RIG-I、MDA5介导的信号通路发挥着相同的作用。在抗鸭病毒性肠炎(duck virus enteritis,DVE)试验中,研究发现LGP2对MDA5介导的抗病毒信号通路的调节是呈浓度依赖性的,低浓度的LGP2能够增强MDA5介导的抗病毒作用,而高浓度的LGP2则抑制了MDA5的抗病毒作用(图1:IV)。在牛口蹄疫病毒(foot-and-mouth disease virus,FMDV)感染中,上述结论同样得到了印证,LGP2在抗FMDV中起着双向调控类似于平衡器一样的作用。在青鱼抗草鱼呼肠病毒(grass carp reovirus,GCRV)和鲤鱼春季病毒血症病毒(spring viremia carp virus,SVCV)研究中发现,LGP2本身对诱导IFNs表达的作用不大,但低浓度的LGP2可以导致MDA5介导的信号通路和抗病毒活性明显增强。小鼠抗脑心肌炎病毒(encephalomyocarditis virus,EMCV)感染试验表明,LGP2在RIG-I、MDA5介导的抗病毒信号通路中发挥正向调节作用。进一步的研究表明,LGP2的ATP酶活性结构域在这种正向调节作用中扮演重要角色。
三、LGP2在适应性免疫应答中的作用
LGP2在天然免疫应答中发挥正、负调节作用的同时,在适应性免疫应答中同样也发挥着重要的调节作用。在适应性免疫应答中,LGP2对CD8+ T细胞的产生、分化、存活与凋亡发挥调控作用。T细胞增殖、存活与发挥效应需要Dicer(一种特定的核糖核酸内切酶)的参与,LGP2能够与Dicer酶相互作用,促进T细胞存活,提高其适应性。
Kandasamy等试验发现,病毒感染宿主细胞时,LGP2可以促进外周T细胞增殖,增强外周T细胞的免疫应答效应,提高机体免疫力。在MAVS-/- CD8+ T细胞试验中进一步发现,抗原刺激机体后,细胞内死亡受体(death receptor)介导的细胞凋亡通路被激活,在无MAVS参与的条件下,LGP2能够抑制该凋亡通路,维持CD8+ T细胞存活。图2是依据文献绘制的LGP2调控CD8+ T细胞的存活与凋亡路径。
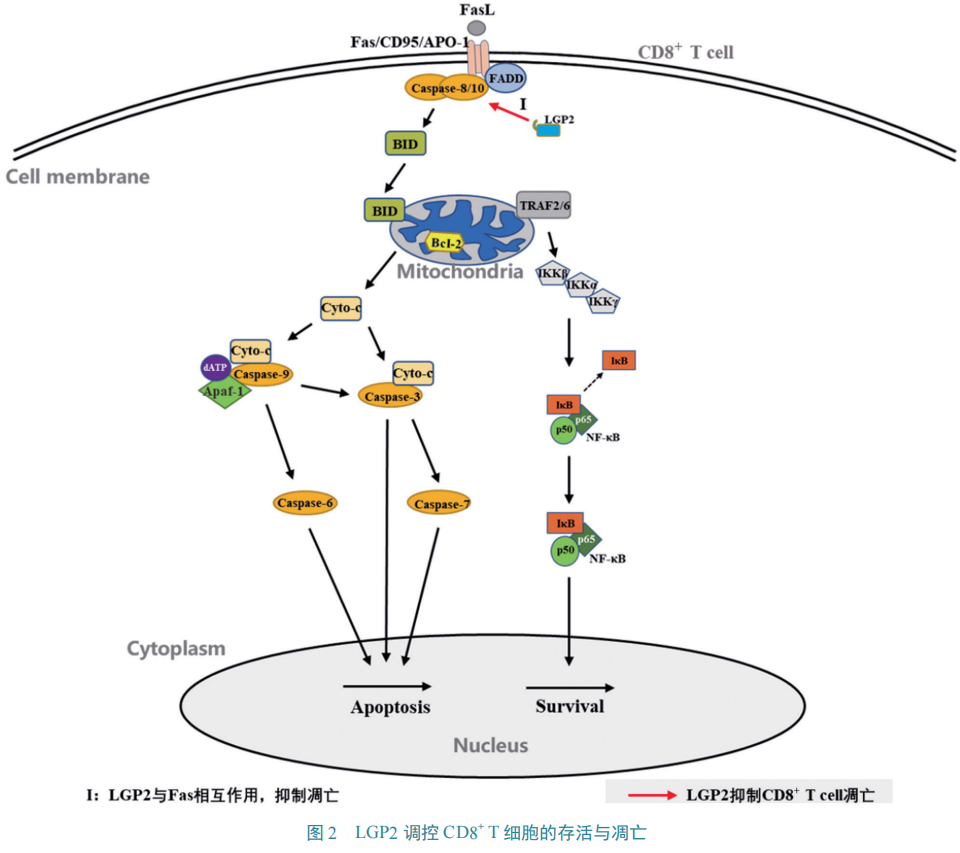
最新研究表明,LGP2可通过两种机制调节CD8+ T细胞凋亡与抗凋亡:(1)激活T细胞受体(T cell receptor,TCR),促进LGP2过表达,而过表达的LGP2通过与细胞表面受体Fas(factor related apoptosis,或称CD95/APO-1)相互作用,延缓CD8+ T细胞的凋亡进程(图2:I),从而维持CD8+ T细胞的正常功能。(2)机体受到抗原刺激后,LGP2能够调节T细胞微小RNA(microRNA或称miRNA)的合成,从而调控促凋亡因子的释放和T细胞的存活。同时,LGP2还能够协助CD8+ T细胞识别多种病毒RNA,清除体内的病原微生物。
参考资料
