【1.21】T细胞协调细胞介导的免疫并调节B细胞对抗原的反应
一些细菌和寄生虫,以及所有病毒,都在细胞内复制,它们无法被抗体检测到,而抗体只能进入血液和细胞外空间。细胞内入侵者的破坏是T淋巴细胞的功能,T淋巴细胞负责细胞介导的适应性免疫应答。但T淋巴细胞参与对多种病原体的反应,包括细胞外生物,因此必须发挥多种效应器活性。
胸腺中发育有几种类型的T淋巴细胞。它们的特征在于它们表达的T细胞受体的类型和某些标记物的表达。两类主要的T细胞表达一种叫做CD8的细胞表面蛋白或另一种叫做CD4的细胞。这些不仅是随机标记,而且对T细胞的功能很重要,因为它们有助于确定T细胞和其他细胞之间的相互作用。回想第1-10节,T细胞检测源自细胞表面MHC分子显示的外来抗原的肽。CD8和CD4通过识别MHC分子的不同区域并参与与其抗原结合的T细胞受体的信号传导,在抗原识别中发挥作用。因此,CD4和CD8被称为共受体(co-receptors),它们提供CD8和CD4T细胞之间的功能差异
重要的是,有两种主要类型的MHC分子,称为MHC类I和MHC类II。它们的结构略有不同,但都在外表面有一个细长的凹槽,可以结合肽(图1.29)。在细胞内MHC分子的合成和组装过程中,肽被困在这个凹槽中,然后肽:MHC复合物被运送到细胞表面并显示给T细胞(图1.30)。因为CD8识别MHC I类蛋白的区域,而CD4识别MHC II类蛋白的区域,这两个共受体在功能上区分T细胞。因此,CD8 T细胞选择性地识别由MHC I类分子呈现的多肽,而CD4 T细胞选择性地识别由MHC II 类分子呈现的多肽
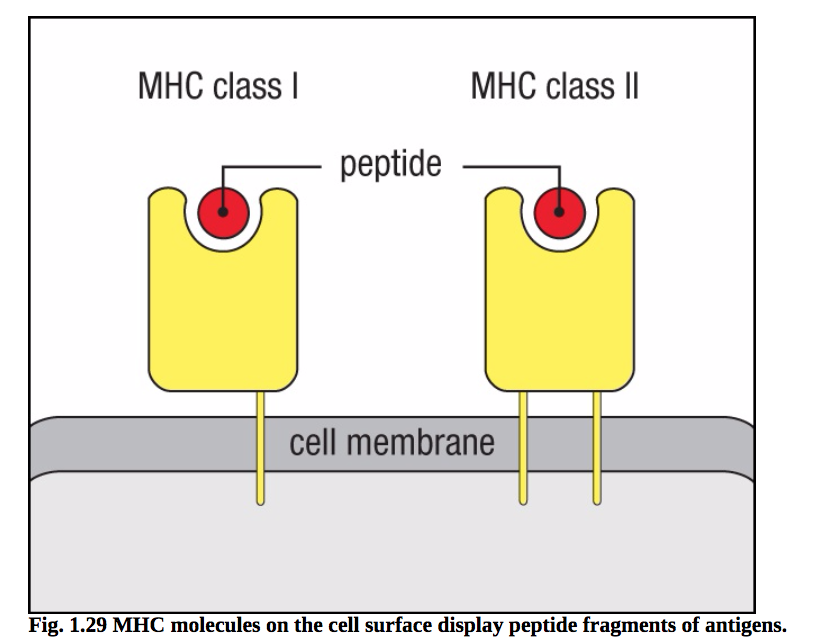
图1.29MHC分子在细胞表面显示抗原的肽片段。MHC分子是膜蛋白,其胞外结构域形成肽片段结合的裂解蛋白。这些片段来源于细胞内降解的蛋白质,包括自身和外来蛋白质抗原。肽在到达细胞表面之前被新合成的MHC分子结合。有两种MHC分子,MHC I类和MHC II类;它们具有相关但不同的结构和功能。虽然为了简单起见,这里没有显示,但MHC I类和MHC II类分子都是两条蛋白质链和结合的自身或非自身肽的三聚体
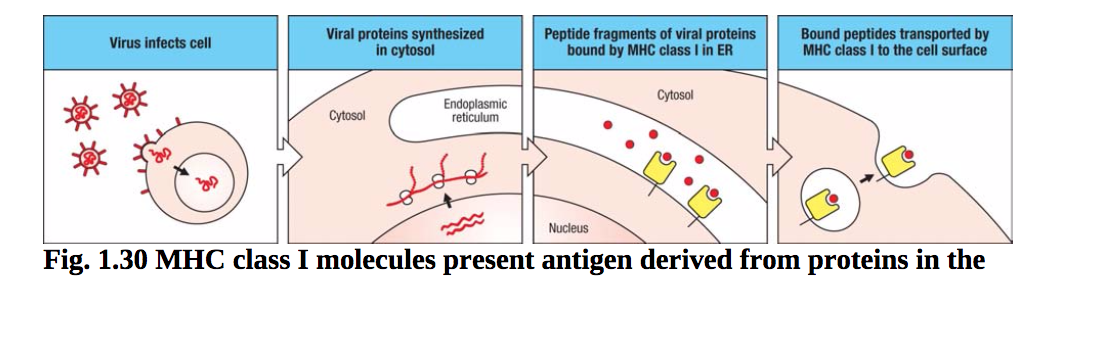
图1.30 MHC I类分子的抗原来源于细胞质中的蛋白质。在被病毒感染的细胞中,病毒蛋白在细胞质中合成。病毒蛋白的肽片段被运送到内质网(ER),在那里它们被MHC I类分子结合,然后将肽运送到细胞表面。
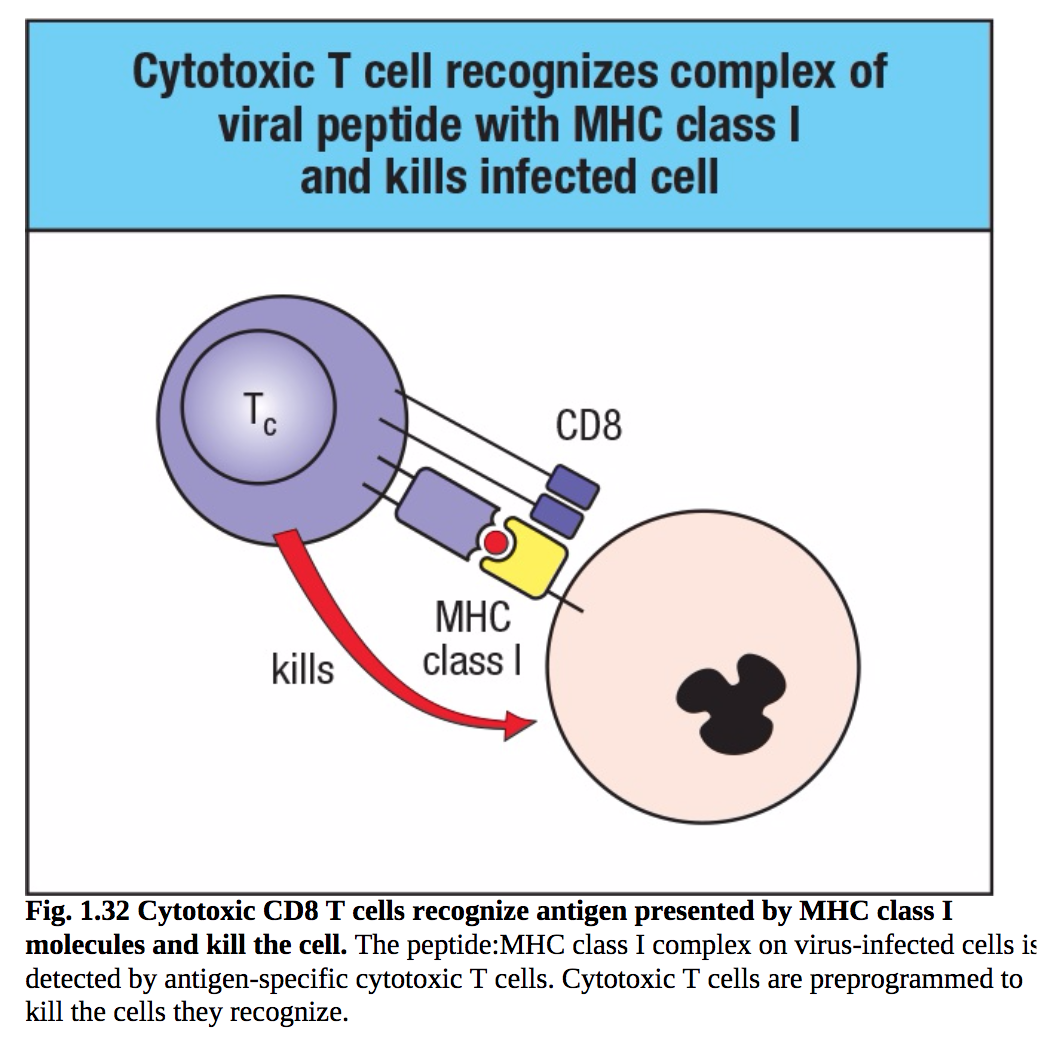
T细胞最直接的作用是细胞毒性。细胞毒性T细胞是对抗感染病毒的细胞的效应T细胞。来自病毒在感染细胞内增殖的抗原显示在细胞表面,在那里它们被细胞毒性T细胞的抗原受体识别。然后,这些T细胞可以通过在病毒复制完成并释放新病毒之前直接杀死受感染的细胞来控制感染(图1.31)。细胞毒性T细胞携带CD8,因此识别MHC I类分子呈现的抗原。由于MHC I级分子在身体的大多数细胞上表达,因此它们是防御病毒感染的重要机制。携带病毒肽的MHC I类分子被携带CD8的细胞毒性T细胞识别,然后杀死受感染的细胞(图1.32)
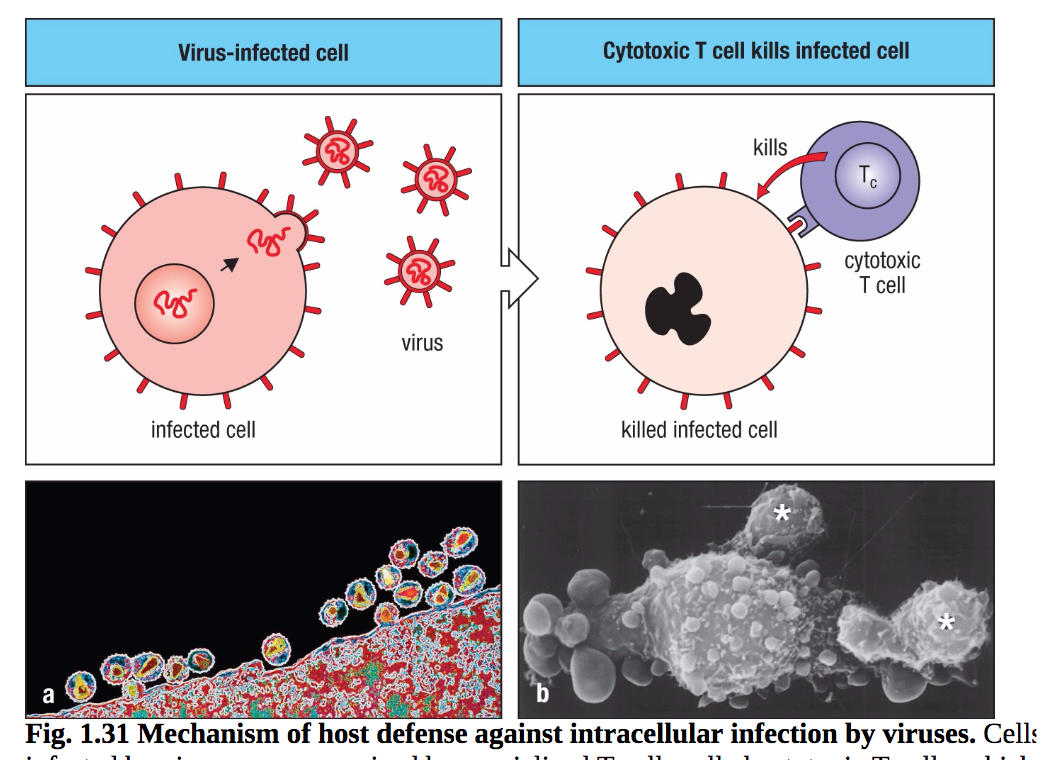
图1.31宿主对细胞内病毒感染的防御机制。被病毒感染的细胞被称为细胞毒性T细胞的特化T细胞识别,并直接杀死被感染的细胞。这种杀死机制涉及到被称为半胱天冬氨酸的酶的激活,这种酶的活性位点含有半胱氨酸,并在天冬氨酸处切割靶蛋白。这些半胱天冬酶反过来激活一种胞浆核酸酶,在被感染细胞中切割宿主和病毒DNA。图a是人类免疫缺陷病毒(HIV)从受感染t细胞表面萌芽的彩色透射电镜图。图b为两个细胞毒性T淋巴细胞杀死靶细胞的扫描电子显微图(星号)。
CD4 T细胞识别MHC II类分子所呈现的抗原,该抗原由免疫系统的主要抗原呈递细胞(树突状细胞、巨噬细胞和B细胞)表达(图1.33)。因此,CD4T细胞倾向于识别细胞外环境中吞噬作用所摄取的抗原。CD4 T细胞是本章前面提到的辅助T细胞。它们发育成多种不同的效应子群,称为TH1(T辅助型1)、TH2、TH17等,它们产生的细胞因子与前面提到的ILC亚群相似,激活保护不同病原体的效应子模块。这些子集主要作用于,外周组织感染或损伤。在淋巴组织中,CD4 T细胞的一个子集,称为T滤泡辅助细胞(TFH),与B细胞相互作用,以调节免疫应答过程中的抗体产生。第9章描述了各种T辅助子集。
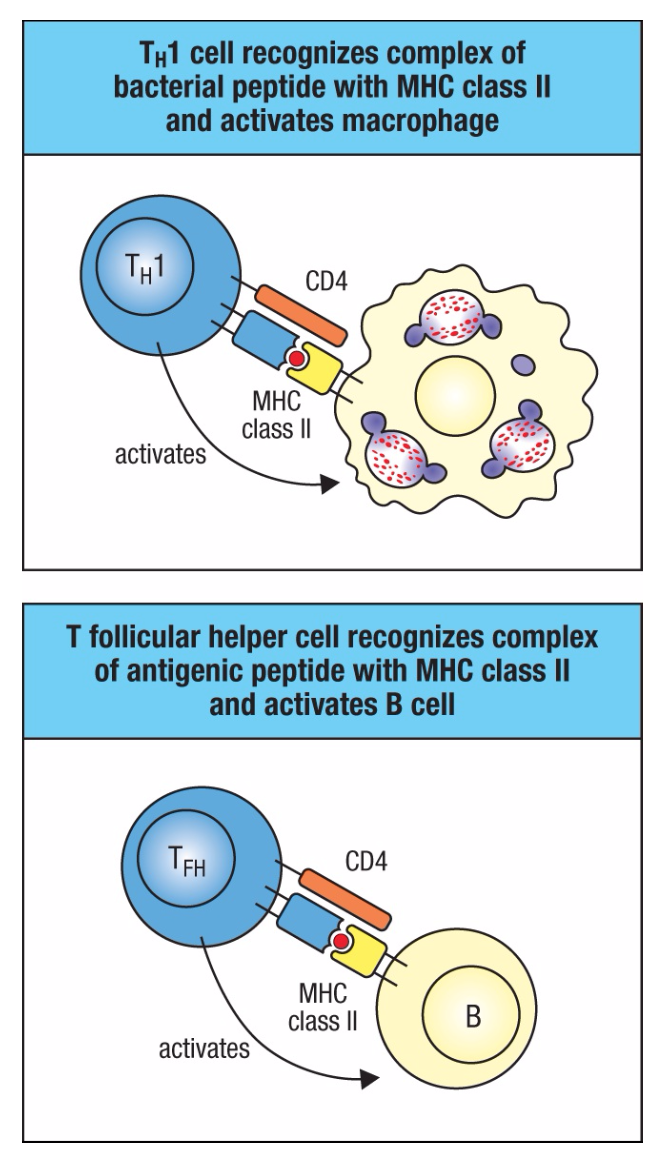
图1.33 CD4 T细胞识别MHC II类分子呈现的抗原。TH1细胞在被感染的巨噬细胞上识别其特异性抗原后,激活巨噬细胞,导致细胞内细菌的破坏(上图)。当滤泡辅助细胞(TFH)识别B细胞上的抗原时(下图),它们激活这些细胞增殖并分化为产生抗体的浆细胞(未显示)
例如,CD4 T细胞的TH1亚群有助于控制巨噬细胞内膜囊泡中的某些细菌。它们产生与ILC1细胞相同的细胞因子IFN-γ,IFN-γ激活巨噬细胞以增加其细胞内的能力并摧毁这些细菌。受该功能控制的重要感染是结核病和麻风病,分别由结核分枝杆菌和麻风分枝杆菌引起。分枝杆菌在细胞内存活,因为它们阻止它们占据的囊泡与溶酶体融合,溶酶体含有多种降解酶和抗微生物物质(图1.34)。然而,在其表面,受感染的巨噬细胞呈现分枝杆菌衍生的抗原,这些抗原可以被激活的抗原特异性TH1细胞识别,其继而分泌特定的细胞因子,诱导巨噬细胞克服囊泡融合的阻滞。TH2和TH17亚群分别产生专门用于促进对寄生虫或细胞外细菌和真菌的反应的细胞因子。CD4 T细胞及其特异性亚群在适应性免疫中发挥着广泛的作用,我们将在本书中多次提及它们,包括第8、9、11和12章
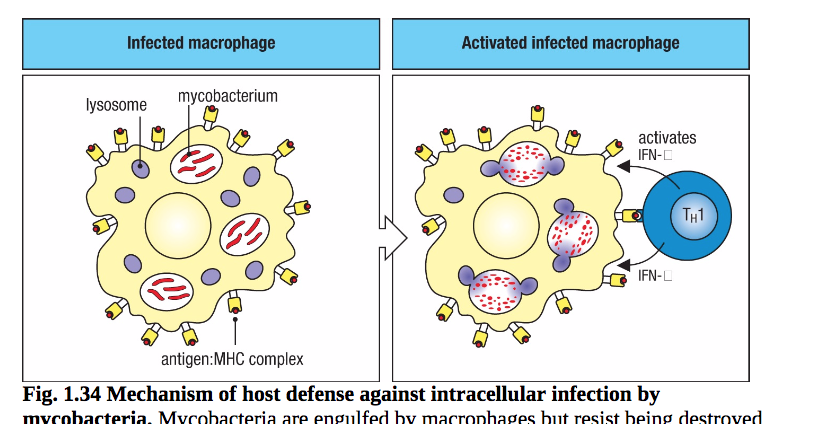
图1.34宿主防御分枝杆菌胞内感染的机制。分枝杆菌被巨噬细胞吞噬,但通过阻止其所在的细胞内小泡与含有溶酶体的杀菌剂融合而抵抗被破坏。这样,细菌就不会被杀死。在巨噬细胞中,分枝杆菌在这些囊泡中持续存在并复制。当,吞噬细胞被TH1细胞识别并激活,然而,吞噬泡与溶酶体融合,细菌可以被杀死。巨噬细胞活化由TH1细胞控制,既避免组织损伤又节省能量
参考资料
- Janeway’s Immunobiology 10th edition
