【8.4.1】延长治疗性蛋白半衰期的当前策略
大分子蛋白质和肽治疗剂已被证明能有效地精确治疗人类的关键疾病。由于生物技术的进步,在过去的几十年中,大量的蛋白质和肽治疗剂进入了制药市场。然而,在临床应用过程中,蛋白质治疗的最大挑战之一是它们在血清中的快速降解以及由于酶促降解,肾脏清除,肝代谢和免疫原性而导致的快速消除,这归因于半衰期短。
- 蛋白质分子的大小和疏水性使其易于肾脏过滤和肝脏代谢。
- 另一方面,负责蛋白质破坏的蛋白酶体具有特异性识别几乎所有种类的外来蛋白质的能力,同时避免了对细胞成分的任何不希望的破坏。
目前,市场上几乎所有基于蛋白质的药物制剂都以高剂量的频率静脉内(IV)或皮下(SC)给药,最终导致剂量波动相关的并发症并大大降低了患者的依从性。因此,通过在蛋白质上附着一个增加总体大小(例如PEG)或帮助受体介导的再循环(例如白蛋白)或操纵氨基酸链的分子,人为地增加蛋白质的治疗半衰期一些更易于避免聚集体形成的革命性方法可以避免体内快速降解。半衰期延长技术能够显着提高循环蛋白的半衰(2–100倍),从而改善其整体药代动力学(PK)参数,已成功应用于激素和酶等多种蛋白质治疗剂中,生长因子,干扰素的凝血因子。综述的重点是评估迄今为止在增强循环半衰期和改善蛋白质治疗功效方面取得的技术进展。
一、前言
肽/蛋白质作为生物治疗剂的引入可能是现代健康科学领域最大的成功之一。自从胰岛素被用作临床上的第一种治疗性蛋白质以来,随着激素,酶,凝血因子和抗体的发展,蛋白质治疗领域迅速发展[1]。目前,许多潜在的肽类药物正在临床研究中。在肽和蛋白质治疗性临床管理中要解决的最大挑战之一是由于蛋白质药物半衰期短而导致的快速降解。由于在全身循环中快速的肾脏清除和明显的酶活性,蛋白质分子通常在血液中的停留时间短[2]。因此,通常需要注射更高剂量以在血清环境中维持治疗浓度达期望的时间段。然而,具有高初始峰的治疗性蛋白质的浓度波动会产生副作用。此外,许多蛋白质的治疗范围非常狭窄。如果可以延长治疗性蛋白质的血浆半衰期,则可以解决这两个问题,因此,以较低的剂量可以维持更长的作用[1]
在过去的几年中,在蛋白质半衰期延长技术的开发方面做出了巨大的努力。 已经提出并测试了许多技术来长效配制治疗性蛋白质,例如氨基酸改变,其他蛋白质(免疫球蛋白结构域或血清蛋白质,例如白蛋白)的遗传融合,与聚合物结合(聚唾液酸化,HESylation,PEG化等) )[3]。 这些技术可分为4个主要类别:
- 氨基酸操纵(amino acid manipulation)
- 生物结合(bio-conjugation)
- 通过将肽连接至聚合物(天然/合成)的翻译后修饰 post-translational modification by attaching the peptide to polymers (natural/synthetic)
- 载体介导的递送。 carrier mediated delivery
前三种技术基于半衰期和循环时间调节,而载体介导的输送概念基于建立某种长效制剂并在循环中以受控和持续的方式缓慢释放治疗剂,从而能够形成长效制剂 。
二、体内蛋白质调节机制和识别“非自身”蛋白质影响蛋白质半衰期的机制
关于蛋白质治疗性给药的主要关注是高消除率。如果蛋白质分子由于折叠错误或损坏而被认为对细胞有毒,则将其从循环系统中移除;还从循环中除去调节蛋白以维持其水平。细胞会去除无法识别为“自身”的任何外来蛋白质。内源性和外源性蛋白质均可因适应性免疫反应而降解[4]。对于细胞蛋白酶体来说,识别外源蛋白质是至关重要的,但也是非常具有挑战性的工作。要选择所有外源蛋白质,细胞蛋白酶体需要技巧。
- 在这里,蛋白酶体不能是序列特异性的,因此任何单个蛋白酶体单元都可以作用于多种不同的蛋白质。
- 另一方面,它必须具有选择性和区分性,以避免任何有害的细胞成分破坏[4]。蛋白酶体如何做到这一点尚不完全清楚。
三、影响循环中蛋白质半衰期的因素
除了具有半周血清半衰期的单克隆抗体外,大多数蛋白质治疗剂都会很快从体内清除[1]。 半衰期短,消除快是蛋白质药代动力学特性的结果[5]。 蛋白质消除的速度取决于几个因素,例如:
- 大小
- 分子量
- 表面电荷[1,6]。
因此,半衰期延长策略的成功取决于该技术是否可以操纵这些因素中的一个或多个。 通过酶促降解,肾脏清除和肝代谢发生蛋白质清除
蛋白质的大小及其疏水性使治疗剂易于肾脏过滤和肝脏代谢。例如,肽的肾脏滤过截留尺寸小于70 kDa。也就是说,小于该肽的肽很容易通过肾脏过滤系统清除[5]。肾脏清除率可通过增加蛋白质大小(流体力学体积)和分子量来降低。
- 一种方法是通过将蛋白质连接到moiety上,例如聚乙二醇(PEG)。
- 另一方面,具有负表面电荷的分子能够通过排斥基底膜的负表面电荷来避免肾脏清除。这使得蛋白质表面修饰成为延长半衰期的另一种选择。
在肝脏代谢过程中,蛋白质被肝细胞摄取。摄取通常是受体介导的。然后,蛋白质分子像其他蛋白质一样,被酶在溶酶体中降解,并从循环中清除。但是,循环中存在内源性蛋白质,可以通过特异性受体介导的再循环来避免肝脏代谢。大多数此类研究集中于白蛋白和IgG的FcRn受体偶联回收。并且观察到,治疗性蛋白质与IgG的白蛋白或Fc部分的结合有助于靶蛋白像其部分一样再循环回到循环中。使用白蛋白缀合设计的第一个半衰期延长的蛋白质疗法之一是长效胰岛素类似物胰岛素德米尔(6)。图1是通过避免肾脏清除和受体介导的再循环的半衰期延长策略的示意图。
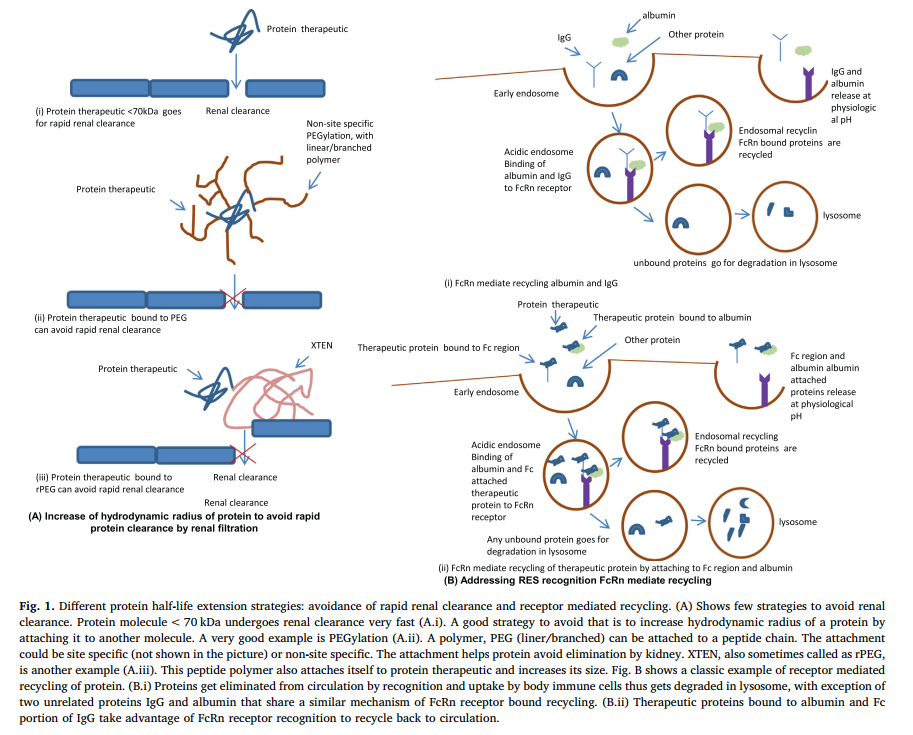
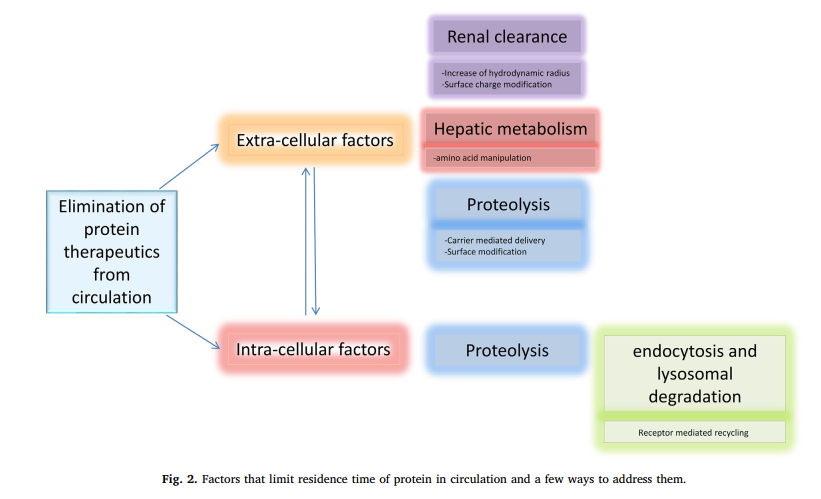
酶促降解可能是决定蛋白质半衰期的最大因素,它可以是位点特异性的或非位点特异性的,发生在身体的任何部位,与任何特定器官无关。也可以是细胞内或细胞外的。酶促降解取决于蛋白质的大小以及二级和三级结构。糖基化水平也影响该过程。治疗性蛋白质的细胞外降解与内源性或膳食蛋白质的降解相同,而细胞内降解通常发生在溶酶体中[6]。防止蛋白质在GIT或其他器官中发生细胞外降解的几种策略之一是借助部分(如PEG)掩盖靶蛋白,或将其掺入由可保护药物的复杂药物输送系统制成的覆盖物或基质中酶促作用。一个例子是修饰carbon介导的胰岛素传递尝试[7,8]。图2总结了限制蛋白质在循环中停留时间的机制,并仅列举几个例子来解决这些问题。
四、克服蛋白质治疗药物体内不稳定性的方法
已经发现许多策略可有效延长治疗性蛋白质和肽的半衰期。这些技术分为以下几类:
- 已证明氨基酸操纵-肽链中一个或几个氨基酸的插入/缺失或改变可降低体内免疫原性和蛋白水解的不稳定性[3]。
- 生物结合-肽与血清蛋白(如白蛋白或免疫球蛋白)结合。基因工程方法也可以用于生产这种融合蛋白[3,5]。
- 翻译后修饰-将肽连接至聚合物(天然/合成)。一个很好的例子是聚乙二醇化。
- 载体介导的递送-在体内环境中附着载体以保护肽分子。一些载体能够在血清中缓慢和持续释放治疗剂。载体可以是微球,脂质体,基质和微米/纳米颗粒[3]。
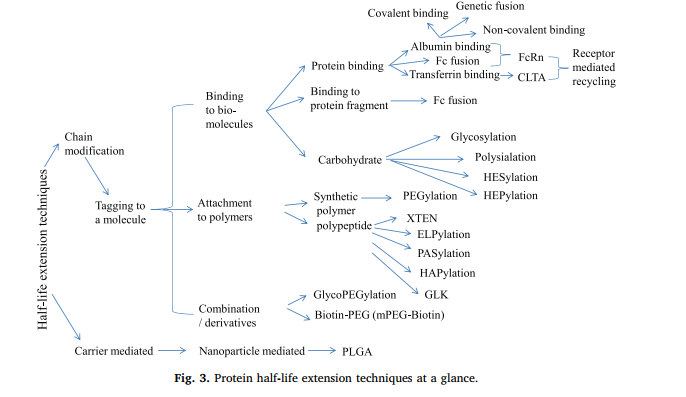
多年来,基于这些策略出现了许多技术。图3总结了迄今为止报道的蛋白质半衰期延长策略。
五、将治疗性蛋白质/肽附着到聚合物上
5.1 聚乙二醇化 PEGylation
聚乙二醇化是一种非常流行的蛋白质修饰技术,于70年代初被引入[9-11]。 肽链共价连接至PEG的一个或多个高分子量链[1],PEG是由环氧乙烷(CH2eCH2eO)的重复单元构成的分子[12]。 该延长通过隐藏肽的抗原决定簇使抗体无法将其识别为异物,并增加蛋白质大小,从而使其不易被肾脏清除[1,11]。 PEG本身是一种不带电荷的亲水性聚合物,它是不可生物降解的,大部分是无毒和非免疫原性的。 高度灵活的PEG分子可轻松连接至治疗性分子[5]。 聚乙二醇化还通过使蛋白质更易溶于水来帮助吸收增强,并掩盖了分子免受蛋白水解降解的作用[11]。
第一代PEG化使用与肽连接的PEG线性链,而第二代提出了支链PEG和PEG衍生物连接的想法[12]。多年来,开发出了更好的PEG化策略,以减少肽的非特异性活性并提高其稳定性。市场上已经有超过10种肽疗法,目前正在临床试验中基于这种技术设计许多潜在药物[13]。弗兰克·戴维斯(Frank Davis)和他的同事在20世纪70年代的第一批作品产生了第一种商品化的聚乙二醇化蛋白药物,即修饰的腺苷脱氨酶(ADA)[11,14,15]。酶腺苷脱氨酶(ADA)主要用于治疗年轻人中的严重合并免疫缺陷病(SCID)[15,16]。像大多数酶一样,ADA的血浆半衰期非常短,只有几分钟。将PEG与牛ADA缀合可将其半衰期延长至小鼠24小时。在随后的研究中,通过肌内(IM)注射对两个孩子进行了PEG-ADA测试。发现该酶从注射部位到血浆迅速吸收,显示出48-72小时的延长半衰期。每周一次,以15 U / kg体重的剂量给药,可以成功维持血浆ADA活性,比正常受试者的红细胞ADA活性高2-3倍[11,15]
Filgrastim是一种重组蛋氨酸人类粒细胞集落刺激因子(G-CSF)(rmetHuG-CSF),在化疗期间需要刺激中性粒细胞的供应来预防发热性中性粒细胞减少症(FN)。以前,它是作为日常注射给药的。非格司亭的聚乙二醇化形式是非格司亭,每个周期仅需给药一次[17,18]。在动物研究过程中,培格非司亭在正常和中性粒细胞减少症小鼠中均能增强和维持中性粒细胞计数。在正常小鼠中单剂量的培格非司亭可将中性粒细胞计数提高至15.5×109 / L。该结果等同于非格司亭的常规剂量,每天两次,持续4天。中性粒细胞的升高水平维持5天。同样,在嗜中性白血球减少的小鼠和骨髓抑制的恒河猴中,嗜中性粒细胞水平升高[19]。由于PEG部分,培格非司亭具有增加的大小,并且不容易被肾脏清除。培格非司亭的血清半衰期延长了42小时,而非格司亭的半衰期延长了3.5–3.8小时[17]
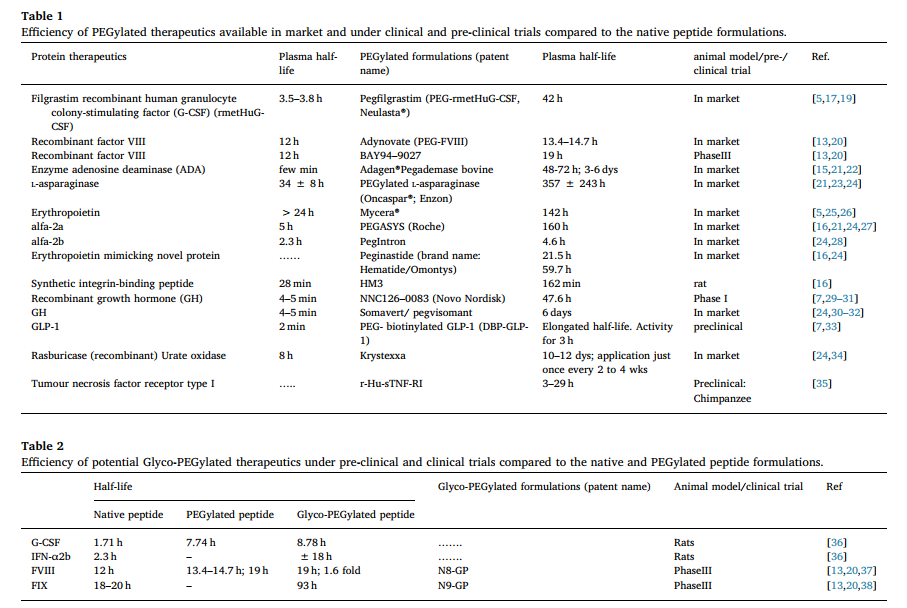
使用聚乙二醇化技术的另一个成功案例是持久的血友病药物。 由于凝血因子缺乏,血友病是一种罕见的血液病。 因此,用重组VIII因子产物处理。 市售产品的半衰期为12小时,每周至少需要注射3次[20]。 FDA批准的聚乙二醇化抗血友病药Adynovate(PEG-FVIII)将半衰期延长至13.4-14.7小时[20]。 同样,采用PEG化技术的III期临床产品(如来自Bayer’s的BAY94-9027)的半衰期为19小时。 Baxalta的Bax855也是如此。 表1提供了市场上可得到的和临床试验中的PEG结合治疗剂清单。
聚乙二醇化被认为是最流行的蛋白质半衰期延长技术。 FDA承认PEG为GRAS(通常被认为是安全的,generally recognized as safe)。 然而,PEG不是100%安全的,因为它是不可生物降解的,具有免疫原性的,并且在肾皮质肾小管上皮细胞中显示出液泡[5]。
5.2 修饰的PEG化 Modified PEGylation
几十年来,人们一直在努力通过将PEG与其他部分修饰和结合来进一步开发PEG结合治疗剂。 将PEG与另一个分子缀合是相对较新的方法。 到目前为止,尚未将任何产品商业化,但是有几份报告显示了它们的潜力。
5.2.1 糖基PEG化 Glyco-PEGylation
常规地,PEG通过与氨基酸上的反应性基团连接而与蛋白质共价连接。 新的“糖基PEG化”方法将PEG连接到O-聚糖上。 因此,它是糖基转移酶的“定点PEG化”。 该方法声称可以最大程度地减少同工型(isoform)形成,并提高半衰期时的PEG性能[36]。 缀合了不同的治疗性肽,并显示可增加循环半衰期[36]。 表2总结了与天然和聚乙二醇化的对应物相比,聚乙二醇化的制剂的效率。
5.2.2 生物素化PEG
PEG的修饰形式是最近通过将PEG与抗生蛋白链菌素结合而开发的[39]。 将生物素化的PEG用于载有吉西他滨的碳磷灰石纳米颗粒( carbon apatite nanoparticle,Co3AP NP)的表面改性。 在小鼠模型中,发现生物素化的PEG可延长Co3AP-吉西他滨复合物在血液循环中的停留时间[39]。 早些时候,在另一项研究中,S Ke,John C. Wright和G S. Kwon(2007)使用牛羧肽酶A(CPA)作为目标酶,并使用不同分子量的PEG成功制备了几种不同的亲和素-生物素-PEG-CPA复合物制剂。 与抗生物素蛋白结合。 体外结果显示,半衰期为3-4天(取决于PEG链长),表明循环停留时间有很好的改善[40]。 因此,它作为蛋白质和肽治疗剂的药代动力学增强剂具有巨大的潜力。
六、将治疗性蛋白质/肽连接到重组多肽
近年来,出现了新的蛋白质半衰期延长技术,该技术包括非结构化多肽链。 就像PEG一样,多肽聚合物与蛋白质治疗剂的结合会增加其流体动力学半径,从而使其不易于肾脏过滤。 但与PEG不同,肽聚合物替代品是可生物降解,无毒,无免疫原性,亲水性和中性的聚合物,由于其简单的合成技术,它们也具有很高的性价比[5,41]。
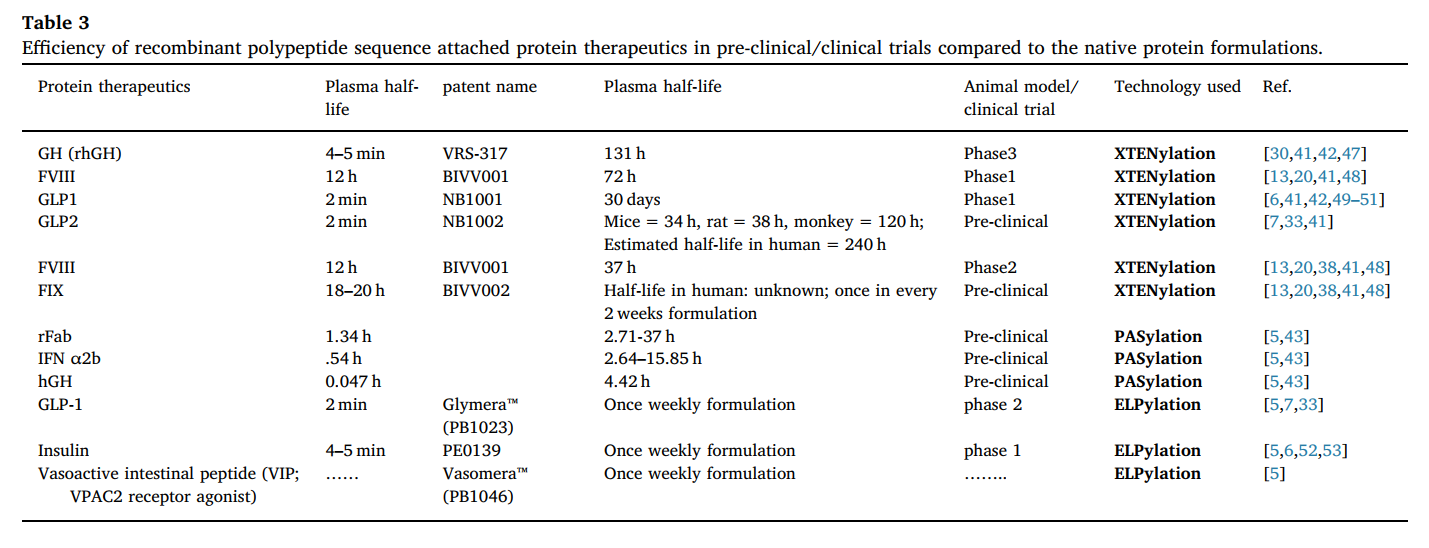
6.1. XTEN
XTEN是Amunix Versatis开发的蛋白质聚合物,可替代不可生物降解的聚合物以延长半衰期。 XTEN是由有限数量的单体组成的非结构化多肽。 与化学聚合物不同,XTEN没有定义的结构(无规卷曲)。 与PEG一样,XTEN也可以通过增加分子的大小和流体动力学半径来增加蛋白质半衰期。 换句话说,XTEN是PEG(也称为rPEG)的良好替代品。 XTEN包含使用大肠杆菌表达的具有A,E,G,P,S和T的氨基酸序列库。 XTEN序列已经过测试,具有增强的半衰期和降低的免疫原性的几种蛋白质疗法[5,42]。 根据Amunix管道产品清单,至少有4种XTEN偶联蛋白治疗剂处于临床试验的不同阶段[42]
6.2 PASylation
PAS是由慕尼黑工业大学开发的另一种肽聚合物,它也像PEG一样工作,并具有可生物降解的肽分子的所有特性。 “ PAS”代表Porline,Alanine和Serine是由100-200个重复序列组成的非结构化肽聚合物,是通过随机排列这三个氨基酸作为结构单元而创建的[5,43]。 最初测试了3种蛋白质作为模型治疗剂:重组Fab片段,IFNα2b和人类生长因子(生长激素)。 连接到Fab片段的不同PAS序列的半衰期增加了2-25倍。 显示PASylated形式的半衰期为2.71-37小时,而rFab本身只有1.34小时。 同样,PASylated干扰素显示血浆半衰期增加5-30倍,hGH显示增加94倍。 XL-Protein GmbH正在研发中的PASylated蛋白疗法有6种,其中大多数处于临床前试验阶段[43]
6.3 ELPylation
另一种可用的多肽融合技术是ELPylation。 像XTEN和PAS一样,ELP(弹性蛋白样多肽)也是包含V-P-G-x-G重复序列的随机序列,主要存在于弹性蛋白中,其中x代表脯氨酸以外的任何蛋白质。 序列与弹性蛋白具有高度相似性,使其易于产生人类弹性蛋白酶,赋予其生物可降解特性。 ELP可以在基因水平上连接到任何肽治疗剂上,并产生热响应性可逆相变。 因此,在一定的阈值温度以上,它们通过形成聚集体而从溶液中沉淀出来。 然而,就像PEG和其他多肽制剂一样,ELP通过增加流体动力学半径而起到半衰期延长剂的作用[5,44]。
6.4 HAP化
富含甘氨酸(Gly4Ser)n多肽的重复序列是HAPylation的活性分子,它由100-200的重复残基组成。 在动物模型中,HAPylation显示可增加Fab的半衰期[5]。 200个残基的HAP共轭Fab在体内的半衰期延长了6小时。 (Gly4Ser)n溶解度遇到重大挫折。 HAP的溶解度取决于链长,长度增加会大大降低溶解度。 较短链的效率不足以延长半衰期[41]
6.5 明胶样蛋白(GLK)Gelatin-like protein
明胶样蛋白(GLK)是一组具有生物活性的重组多肽,具有延长的体内半衰期,并具有(GlyXY)n结构,其中X和Y表示除Cys外的任何天然氨基酸,n介于60至1500之间 氨基酸残基(优选200–1000)质量为6–150 kDa(优选20–80 kDa)[45]。 通过基因融合制备的不同GLK制剂在大鼠中将G CSF半衰期从1.76h增加至10h。 另一方面,琥珀酰化明胶与大豆胰蛋白酶抑制剂和铜/锌超氧化物歧化酶的化学偶联显示半衰期增加了6-7倍[5,46](表3)。
七、将治疗性蛋白质/肽附着到具有生物活性的天然蛋白质或蛋白质结构域(蛋白质片段)上
很少有天然蛋白质分子具有超长的循环半衰期。因此,像血清白蛋白和转铁蛋白这样的蛋白质首先被认为是其他治疗性蛋白质的半衰期延长部分。转铁蛋白的半衰期长是由于独立于转铁蛋白受体,而白蛋白的半衰期长是新生的Fc受体(FcRn)依赖循环的结果。 pH依赖的FcRn受体介导的回收利用的相同机制也适用于人免疫球蛋白(Ig)G抗体。人IgG1,IgG2和IgG4的体内半衰期为14-21天。免疫球蛋白分子的一个片段(Fc区)也被证明是成功的半衰期延长剂[5]。在结构和生物学上,IgG和白蛋白没有任何相似性。但是,它们几乎没有共同的特征。两种蛋白质都有明显更长的半衰期,与血清浓度成反比。这些特征归因于与FcRn受体的结合以及随后的转胞吞作用和回收机制[54]。
7.1 白蛋白结合 Albumin binding
白蛋白具有一些独特的功能,例如〜19 d的血浆半衰期和带负电荷的表面积的多价结合位点,使该分子成为药物输送载体和肽类药物半衰期延长剂的理想候选者。 肽很容易通过共价键和非共价键与白蛋白结合[52,53]。 白蛋白与肽的结合可以通过一种以上的方式并借助不同种类的辅助分子来促进,例如抗体片段(Fab,scFv,结构域抗体(dAbs),纳米抗体),支架蛋白,细菌白蛋白结合结构域,脂肪 酸和其他肽[6,52,53]。 图4和图5是白蛋白与治疗性蛋白质结合的不同模式及其在体内的作用机理的简化图示。

7.1.1 白蛋白的非共价结合 Non-covalent binding of albumin
间接结合可以通过在辅助分子的帮助下将白蛋白与肽治疗剂结合来实现,例如,通过非共价结合将脂肪链连接起来。白蛋白是一种经过充分研究的转运蛋白,具有足够的效率,可逆地结合一系列内源性配体,例如脂肪酸,胆红素和少量外源性配体,例如青霉素,华法林和diazepam [6,52,53]。使用脂肪酸作为配体也称为“脂化” [55]。与这些分子结合的白蛋白增强了它们的生物利用度。利用该技术开发的两种非常著名的肽治疗剂是Levemir®(胰岛素detemir)和Victoza®(利拉鲁肽)。地特胰岛素是通过将胰岛素分子连接到脂肪酸(肉豆蔻酸)上而开发的。胰高血糖素样肽-1激动剂(GLP-1)(来拉鲁肽)也可以通过将脂肪酸(肉豆蔻酸)附着在N端赖氨酸上来产生。一旦制剂进入血流,脂肪酸就与循环中存在的白蛋白形成非共价结合,然后缓慢解离,因此,增加了治疗剂在体内的停留时间。两种配方均规定每天使用一次,而不是多次SC给药[6,52,53]。
7.1.2 与白蛋白共价结合
治疗性蛋白质与白蛋白分子的共价结合是增加蛋白质半衰期的另一种方法。 可以通过化学方法将蛋白质与白蛋白结合来完成[6,52]。 蛋白质治疗剂与白蛋白的直接融合提供了灵活性,可通过将其与基因修饰的白蛋白或白蛋白衍生物(而非内源性白蛋白)缀合来进一步延长蛋白质的半衰期。 白蛋白衍生物是具有延长的血清半衰期的重组蛋白[6]。
7.1.3 重组白蛋白
合成血浆白蛋白衍生物是进一步修饰和设计与白蛋白结合的延长的半衰期蛋白质治疗剂的绝佳选择。 重组白蛋白也是详细检查白蛋白-肽结合的重要来源。 来自酵母的重组人白蛋白与血清白蛋白相同,因此被认为是最安全的[51]。
7.1.4 遗传融合至白蛋白 Genetic fusion to albumin
像共价连接一样,遗传融合技术也可以产生与白蛋白连接的融合蛋白。通过遗传融合,将目标蛋白分子基因与白蛋白基因融合,然后将合成的融合基因转移并在合适的宿主中表达,并且该宿主开始产生合成的融合肽(蛋白+白蛋白)[6,52]。因为制造是在生物体或细胞系内部进行的,并且可以直接收集完全折叠的功能蛋白,所以这提供了更简单的过程。治疗性肽可以在一个末端(白蛋白的N或C末端)或两个末端(双特异性白蛋白融合物)连接至白蛋白[51]。就结合白蛋白的药物开发而言,该技术已被证明非常流行和成功。在过去的二十年中,许多重组蛋白疗法进入了制药市场。这些包括肽(GLP-1),激素(胰岛素),生长因子(G-CSF)和凝血因子。表4列出了处于临床试验或市场中的蛋白质治疗剂

7.2 Fc融合
最近对Fc融合蛋白的评论估计有约350种基于1gG的蛋白治疗剂正在药物开发中或已经批准并在市场上可用[66]。与我们拥有的任何其他可用蛋白质延伸技术相比,该数字是最大的。 IgG偶联蛋白技术的一个非常重要的里程碑是杂交瘤技术的发明[66,67],该技术可快速生产嵌合抗体和人源化抗体,从而大大降低免疫学方面的关注并改善PK和PD特性[66]。基于IgG的延长的半衰期蛋白主要基于Fc域融合偶联。结构上,免疫球蛋白G(IgG)的恒定区Fc区是二价的,为药物结合提供了灵活性。从概念上讲,二价结合是亲合力效应的另一个好处。可以将蛋白质分子添加到Fc区融合位点(N端或C端)或插入环形成中[68,69]。除了延长半衰期,Fc融合有时还显示出改善蛋白质的生理特性,例如增加溶解度和稳定性[68]。
Fc介导的半衰期延长制剂几乎没有什么值得关注的。 它们是ADCC(抗体依赖性细胞毒性)和CDC(补体依赖性细胞毒性)。 CH3eCH3异二聚化已大大解决了这一问题[68,69]。
假设,白蛋白或Fc融合蛋白遵循其结合物延长的半衰期。 然而,令人惊讶的是,发现很少的Fc融合蛋白的半衰期比其Fc对应蛋白的半衰期略短。 Etanercept,abatacept和alefacept是很好的例子。 似乎蛋白质结合会以某种方式搅动Fc区,从而改变其对FcRn受体的亲和力 [66,68–70](表5和表6)

7.3 转铁蛋白融合 Transferrin fusion
血清转铁蛋白具有众所周知的将铁分子可逆结合和转移至组织的功能。 血清半衰期长为7-10天(未糖基化的Tf为14-17天),转铁蛋白是蛋白质半衰期延长缀合物的良好候选者[5,85]。 与非糖基化转铁蛋白(BRX-0585)偶联的GLP-1或exendin-4(EX-4)的半衰期(约2天)增加了约1000倍。 不幸的是,由于副作用高(脉搏和血压升高),批准的制剂被排除在临床试验之外[85]。 另一项研究尝试了一种新的胰岛素原转铁蛋白(ProINS-Tf)融合蛋白。 ProINS-Tf融合蛋白在小鼠中显示出缓慢的持续降血糖作用,并具有更长的半衰期。 与游离ProINS相比,SC给药后缓慢吸收tmax为5.5 h,后者的tmax为0.3 h。 ProINS-Tf的血清半衰期为7.29小时,而ProINS的半衰期仅为0.5小时[5,86]
八、将治疗性蛋白质/肽附着到碳水化合物上
在生物系统中,蛋白质的稳定和防止酶促降解是通过糖基化实现的,糖基化是蛋白质与碳水化合物的共价修饰[87]。 蛋白质的内在特性会因其链上附着的糖分子而改变,最终改变其四级结构的稳定性和对蛋白酶的抵抗力[76]。 可以操纵这种自然保护系统来设计新的半衰期延长的生物疗法[87]。 此外,寡糖卤化物部分通过向蛋白质添加负电荷来增加大小和降低肾脏清除率的能力使糖基化成为增加治疗半衰期的流行方法。 O链寡糖也可通过阻止其进入蛋白质核心来保护嵌合蛋白质免受蛋白酶降解[88]。
8.1 糖基化
糖基化是天然共翻译或翻译后修饰的一种方式,在内质网和高尔基体中进行,其中碳水化合物部分共价连接于天冬酰胺残基(N糖基化)和苏氨酸/丝氨酸残基(O糖基化)。 成分糖的类型和数量,长度,分支和其他特性取决于在其中产生重组蛋白的表达系统,从而对蛋白的三维结构,药代动力学,效率,免疫原性和半衰期发挥作用。
转基因的甲基营养型酵母巴斯德毕赤酵母(Pichia pastoris)几乎不产生与人一样的糖基化蛋白质,例如EPO [89]。 但是重组蛋白是在哺乳动物系统即CHO细胞中产生的,最近又在人细胞系中产生,因为它们在糖基化方面与自然人衍生的对应物相似[90]。
用这种技术制备的第一代重组蛋白通常带有与天然形式相似的聚糖。 例如,人绒毛膜促性腺激素β亚基的羧基末端区域(C末端肽或CTP)的28个氨基酸长序列的融合增加了某些激素的半衰期。 CTP的编码序列与激素-EPO,FSHβ和TSHβ的N末端编码序列以及GH的C末端和N末端连接。 与TSH-wt相比,在COS-7细胞(非洲绿猴)中表达的TSH-CTP显示出延长的半衰期。 与天然肽(5小时)相比,EPO-CTP的半衰期增加了2-3倍。 FSH-CTP于2010年获得批准(商标名称ELONVA)。 嵌合GH(CTP-GH-CTP-CTP)的半衰期比未糖基化的天然形式(12分钟)长4-5倍[88]。 图6是位点特异性诱变的简化图示。
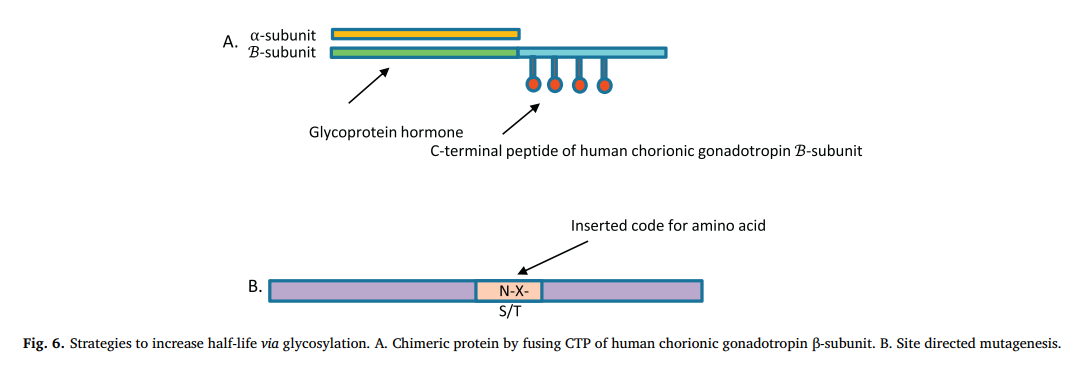
在不中断受体结合和催化位点的情况下对蛋白质编码DNA进行定点诱变是通过糖基化增加半衰期的普遍策略。 插入(N-X-S / T)码,其中X是脯氨酸以外的任何氨基酸,会引入N糖基化位点。 市售的EPO类似物,Darbapoietin alfa带有2种额外的N-聚糖,半衰期为26.3 h,比天然的高3倍[88,91]。 与天然形式相比,在人类生长激素中添加3个额外的N个糖基化位点(N-X-S / T)可将其半衰期延长1至24倍。 但是,效率和N-聚糖插入数量之间的关系并不总是线性的,而是成正比的。 与天然蛋白的0.23 h相比,重组蛋白的重唾液酸化形式将半衰期提高到5.6小时,而中等唾液酸化的3种额外的N联寡核苷酸形式则表现出最佳的IGF-1反应,半衰期为3.10 h [ 92]
Mpl配体是一种造血生长因子,属于糖酵解激素家族,仅包含O联糖基化。 另一方面,饱腹感激素根本不包含聚糖。 S. Elliott等人通过定点诱变分别向这些激素添加了4个和5个N-聚糖,这在小鼠模型中均产生了更高的效率和更长的循环时间[93]。
糖基化对于蛋白质的功能和效率非常重要。 重要蛋白质的任何异常糖基化都可能导致病理状况[94],在使用该技术制备任何新的治疗制剂时必须考虑这些状况。
8.2 聚唾液酸化 Polysialylation
天然存在的PSA(或椰油酸; CA)聚合物是可生物降解的,亲水性高的碳水化合物链,在体内没有确定的受体。因此,用末端唾液酸对治疗性肽和蛋白质进行翻译后修饰可改善药物的药代动力学并降低免疫原性,从而提高其整体效率。据报道,某些细菌利用其聚唾液酸涂层来逃避宿主免疫系统的识别[95],这与聚合物的低免疫原性有关。费尔南德斯等。证明较短的PSA(分子量为22 kDa或更少)与抗癌剂,天冬酰胺酶结合用于治疗白血病增加了酶的稳定性和血清半衰期,并显着提高了天冬酰胺酶曲线下的面积[96]。 Jain等人发表的另一项研究。结果表明,聚唾液酸化胰岛素可显着提高糖尿病的治疗价值。他们发现,与体内完整的胰岛素治疗相比,用22-kDa PSA进行多唾液酸化的胰岛素可以更长时间地降低血糖水平[95]。此外,包含较长的PSA(例如平均分子量为60 kDa)大大延长了小模型药物荧光素的循环时间,该药物的半衰期非常短[97]。使用其他几种蛋白质治疗剂,如抑肽酶和IgG免疫球蛋白,也获得了相似的结果[98]。迄今为止,PSA作为一种有前途的“隐身聚合物”可以通过化学或化学酶策略在体外与蛋白药物偶联。然而,研究的进展也为在大肠杆菌的细胞质中重组蛋白的位点特异性多唾液酸化开发了一种生物合成途径[99]。
8.3 其他基于多糖的方法 Other polysaccharide based approaches
除了流行的技术(例如糖基化和聚唾液酸化)外,在最近的20年中,几乎没有其他方法可以使用。 他们中的大多数都取得了有限的成功,但是其中一些制剂有希望用于临床试验。 然而,它们都显示出作为半衰期延长剂的潜力。
8.3.1 葡聚糖结合 Dextran conjugation
葡聚糖聚合物是具有α(1-3)键的D-葡萄糖聚合物。 葡聚糖和葡聚糖衍生物通过几种不同的方法化学结合到蛋白质上,包括高碘酸盐氧化以及通过酶促方法。 像PEG一样,右旋糖酐也增加了蛋白质的流体动力学半径并减少了肾脏滤过。 作为化合物,它被FDA认可为GRAS。 据报道,葡聚糖偶联的生长抑素正在临床试验中[55]。 其他一些报道提到葡聚糖偶联蛋白,例如尿酸酶[100],超氧化物歧化酶[101],血红蛋白[102],胰岛素[103],血红蛋白[104,105],天冬酰胺酶[106],羧肽酶G2 [107,108]。 不幸的是,这种方法在2006年之后进展不大
8.3.2 羟乙基化 HESylation
植物多糖支链淀粉的修饰形式是羟乙基淀粉(HES),其基本上是天然聚合物。 HES通过支链淀粉的羟乙基化产生。 羟乙基化使多糖更具水溶性,并防止了由于α-淀粉酶的降解[55]。 HESylation技术在包括促红细胞生成素和FVIII在内的几种蛋白质治疗剂的半衰期延长中显示出令人鼓舞的成功[55]。 但是,最近的研究表明,重症患者的HES与肾衰竭和死亡风险增加有关[109]。 另外,由于研究不当,早期的临床试验报告也很少被撤消[110]。 总体安全问题使HESylation方法存在疑问。
8.3.3 羟化 HEPylation
肝素是HEPylation的活性聚合物,它是一种天然多糖,是肝素的生物合成前体。 由于它与肝素的关系,因此在我们的体内环境中被认为是“自我”,没有任何免疫原性问题。 因此,它被认为是可生物降解的蛋白质半衰期延长部分的一个很好的选择。 简化式为[-4 -N-乙酰基葡糖胺-a 1,4-葡萄糖醛酸-b 1-] n [111]。 肝素及其衍生物作为多元领域药理学潜力的潜力备受关注[111]。 肝素显示血流中半衰期为15h至8天,具体取决于MW和给药途径[112]。
8.3.4. 卤化 HAylation
透明质酸(HA)是具有直链和负电荷的天然多糖(重复的二糖:[b(1-4)-GlcA-b(1-3)-GlcNAc] n)。 它存在于人体的不同组织中。 像其他半衰期延长剂一样,HA与任何肽的结合都可以增加蛋白质的大小,从而降低肾脏清除率。 HA还保留水分。 这种流体动力学特性就像蛋白质分子之间的壁或绝缘层一样工作[55]。
九、通过基因操作延长半衰期
9.1 蛋白酶抗性点突变
一个或多个基因的突变已成功地产生了天然短效肽治疗剂的少数长效对立部分。 肽链中一个或多个氨基酸的改变可以增强蛋白质的PK和PD特性,保持其活性不变。 长效胰岛素制剂就是一个很好的例子(白蛋白结合部分前面提到)
最近,点突变已成为一种增强蛋白质药代动力学的新技术。 单个氨基酸取代/改变可以使分子在体内具有较高的蛋白酶抗性。 Nautilus Biotech提出了两种蛋白质制剂,即干扰素β(IFNß)和生长激素[1]。 IFNß的专利申请是在2006年,到目前为止,尚未公开有关进展的详细信息[116],并且生长激素的制剂已于2017年后从1期试验中终止[117]。 即使进展缓慢且推迟商业化,该技术本身仍然很有希望,可以加以改进。
十、纳米粒子(NP)介导的方法 Nanoparticle (NP) mediated approaches
NP介导的受控药物递送载体的设计具有多种目的,例如将药物靶向特定器官进行靶向递送,或保护治疗剂免受在苛刻环境下的降解。 使用NP介导的药物传递来调节治疗药物的PK和PD是另一种产生长效治疗药物的方法[7]。 药物递送系统在常规意义上不改变治疗性分子的PK和PD。 相反,该概念依赖于将天然或重组肽分子掺入基质或另一分子内,该分子的功能类似于保护性覆盖物。 该系统就像一个“仓库”一样工作,而不是一次引入治疗量,而是将药物缓慢释放到全身循环中,从而形成长效制剂[7]
流行的NP系统是PLGA(聚乳酸共乙醇酸)。 很少有非常有前途的PLGA和修饰的PLGA治疗蛋白制剂(例如,聚乙二醇化PLGA)处于临床前/临床试验阶段。 实例之一是PLGA-PEG-PLGA的可生物降解的可溶性热敏三嵌段共聚物,商品名ReGel TM。 ReGel™在室温下可溶于水,但在体内温度下会形成高度粘稠的凝胶,可维持其结构一个月。 ReGel™已测试了多种治疗性蛋白质。 在临床前研究中,掺有ReGel™的胰岛素[7,118,119]和GLP-1 [7,120]显示出非常好的持续释放曲线。
药物递送载体的另一个很好的例子是胶体囊泡脂质体。 脂质体是模仿双层两亲性细胞膜结构的脂质制剂。 设计了几种制剂,它们的脂质组成和粒径互不相同,因此具有不同的PD和PK特性。 脂质体本身在体内的半衰期非常短(<2小时),但与聚合物部分偶联的脂质体(如PEG)具有延长的半衰期(人体内为45小时),可用于长效蛋白质制剂的制备。 掺入脂质体的凝血因子在临床前测试中显示出成功[69]
尽管从概念上讲非常有前途,但是已知药物输送系统的主要缺点在于制造和给药方面的挑战。 蛋白质的突然释放和不良的装载效率也是常见问题[69]。 Nutropin是一种长效生长激素制剂,已于1999年获得批准。它是PLGA内掺入的人类生长激素制剂,长达一个月。 由于与爆发释放相关的副作用(超过50%的治疗剂仅在前几天释放),该产品于2004年停产[7,29,121]。 该技术的未来在于成功解决相关缺陷[69]。
十一、其他延长半衰期的方法(可生物降解和不可生物降解的聚合物以及化学修饰)
已经提出并报道了几种聚合物作为蛋白质半衰期延长部分并可以替代PEG,这些聚合物大多在体外和动物模型中显示出良好的前景。 尽管这些技术看起来很有希望,但它们仍需要至少几年的研究来评估它们在医药市场上是否具有作为半衰期延长剂的真正潜力。 表7报告了其中一些“尚未流行”的半衰期延长剂及其临床前发现
十二、结论
蛋白治疗的半衰期延长的“生物更好”版本已被证明在所有方面都非常有效。 在过去的几十年中,蛋白质新配方的研究和开发花费了很多。 生物技术的进步为该领域的蓬勃发展做出了巨大贡献。 但是,仍然存在需要解决的挑战。 临床安全性和免疫原性问题一直存在。 蛋白质在体内环境中的不稳定性是免疫原性的一个促成因素。 甚至半衰期延长的制剂也可有助于免疫原性。 如果在持续释放过程中不能保持蛋白质的稳定性,则产生的颗粒可以诱导免疫反应,这甚至可能是影响蛋白质稳定性的另一个因素[125]。 简而言之,必须对任何半衰期延长的蛋白质制剂进行广泛的PK和PD研究,以避免所有缺点并消除任何不良副作用
参考资料
- 2019,Current strategies in extending half-lives of therapeutic proteins
