【9.3.7】mRNA体外酶连polyA
一种生成和维持 mRNA体外转录所需的多聚 (A) 编码 DNA 序列的简便方法
sam点评:
- 这种通过酶连的方法得到的polyA,有长有端,怎么评价去掉短的,保证纯度呢?
- ScaI到底发挥了什么作用?
- 2n-8 , 没轮酶切和酶连之后,下一轮序列的长度。
在体外生成 mRNA以编码治疗性或细胞修饰蛋白正在迅速获得青睐。决定体外转录 mRNA翻译效率的一个重要因素是 3' poly(A) 序列的长度。然而,从环状质粒可重复地生成和维护模板以具有一致长度的均聚 (A) 序列是具有挑战性的。此处报告的程序需要使用 IIS 型酶重复限制性消化、连接和循环质粒繁殖。使用该方法产生的大约 100 bp 的均聚序列大约等于哺乳动物细胞 mRNA 中发现的 3' A 残基的数量。
确保 poly(A) 尾的最佳长度对于有效翻译至关重要,而这些序列的缩短会导致 mRNA 衰减(在 [ 3 , 4 ] 中进行了综述)。可以使用聚 (A) 聚合酶将多个 A 残基添加到合成 RNA大肠杆菌(EPAP) [ 5 ] 或在从编码模板转录期间 [ 6 , 7 ]。
-
当使用 EPAP 将 A 残基添加到 mRNA 的 3' 末端时,实现尾长的一致性可能很困难 [ 6 , 7 ]。
-
使用含聚 (A:T) 的环状质粒的一个并发症是这些载体中的均聚 A:T 碱基对不稳定,并且在细菌复制过程中容易缩短 [ 6 , 8 ]。
-
最近使用线性质粒载体来增加较长聚 (A:T) 束的稳定性 [ 6 ]
-
多聚 (A:T) 编码区的序列包含 13 个 A:T 碱基对,侧翼为Faq I 和Bve I 型 IIS 限制性位点。Bve I 位点位于多聚 (A:T) 编码序列的 5' 端,Faq I 位点位于 3' 端。这些类型的 IIS 限制酶在 poly(A:T) 侧翼区域具有独特的切割位点。识别位点的定向使得酶各自在聚(A:T)序列内不对称切割以产生包含四个T或A突出端的互补粘性末端。
-
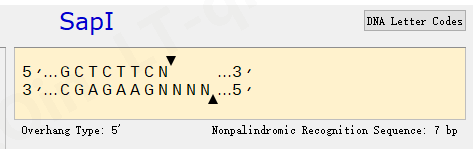
此外,将Sap I 限制性位点置于 poly(A:T) 束的 3' 处,该酶用于线性化质粒以生成模板体外转录。使用定点诱变消除了FLuc ORF 中天然存在的Sap I 位点,同时保留了报告酶的氨基酸序列。
-
延长 A:T 序列过程的第一步需要对质粒进行两次独立的双重消化(图 1B)。用Bve I 和Sca I 或Faq I 和Sca切割后根据酶供应商(New England Biolabs,MA,USA)推荐的程序,纯化了各自包含较长聚(A:T)序列的片段。
-
连接后重建复制质粒(T4 DNA 连接酶,ThermoFisher,MA,USA)并在 XL1-Blue 菌株(Agilent Technologies,CA,USA)的化学感受态细菌中繁殖。ScaI 在氨苄青霉素抗性基因内有一个独特的识别位点,并被选中以尽量减少意外质粒克隆的产生。
-
因此,在该初始消化和连接之后形成的质粒应该含有 18 bp 的 poly(A:T) 序列。重复质粒的限制性消化和连接,并且聚(A:T)道随着每个连续循环而延长。理论上,对于第二到第五个克隆周期,poly(A:T) 序列的长度将分别增加到 28、48、88 和 168 bp。
-
使用标准 Sanger 测序(图 1 C)不可能精确测量 A:T 碱基对的数量,因此同源区的长度是根据包含感兴趣区域的限制性片段的长度来估计的(图 1D)。我们的观察表明,在重复克隆过程的六个循环后,poly(A:T) 片段的最大长度达到了 80 到 100 bp。这类似于 mRNA 中天然存在的 poly(A),应该适用于细胞内翻译 [ 2 ]。
-
在线可用信息表明,这种多聚 (A) 编码序列的长度也超过了合成双链 DNA 的商业供应商通常可以保证的长度。来自克隆反应连续步骤的中间体的线性模板,缺乏FLuc序列,用于使用 T7 RNA 聚合酶(TranscriptAid T7 High Yield Transcription Kit,ThermoFisher)生成 mRNA。mRNA 的电泳分辨率证实了转录物长度的增加,这与 mRNA 的含聚 (A) 的 3' 末端的延长相关(图1E)。

酶切位点信息:



参考资料
- A convenient method to generate and maintain poly(A)-encoding DNA sequences required for in vitro transcription of mRNA。 https://www.future-science.com/doi/epub/10.2144/btn-2018-0120
