【2.1.1.2】自组装mRNA疫苗
mRNA疫苗具有促进抗原瞬时表达(通常是几天)的独特特征。外源性抗原的表达受编码 mRNA 的寿命控制,其受先天降解途径的调节。虽然蛋白质表达的这种短暂性需要重复给药来治疗遗传疾病和癌症,但它对疫苗非常有益,因为初免或初免加强疫苗接种足以在不接触传染病的情况下产生高度特异性的适应性免疫。然而,mRNA疫苗的胜利受到几个因素的阻碍。
- 首先,mRNA 是一种相对脆弱的大分子,容易受到物理、化学和酶促降解的影响。
- mRNA 的这种微妙性质对其储存和治疗的管理施加了限制。
- 其次,抗原的精确鉴定对于改变疾病的进程至关重要。不适当的抗原序列可能会导致免疫系统的意外激活 [ 3 ]。
- 最后,裸露的IVT mRNA通过身体的生理屏障进入靶细胞的能力有限。
因此,使用裸 mRNA 进行治疗可能需要频繁给药和/或高剂量,因此需要对生物制药开发和患者的高依从性进行大量投资。然而,这些挑战可以通过仔细设计目标mRNA 序列和帮助 mRNA 进入细胞的递送载体来规避。
术语“自组装”是指由非共价相互作用驱动的单个分子自发排列成超分子组装体(supramolecular assembly) [ 8 ]。 这种方法在“智能”材料的开发中极具吸引力,因为单个构建块的特性可以微调结构的特性,并且可以通过简单的组分混合来完成制备。此外,自组装完全依赖于构建块之间的物理相互作用,因此可以应用于小分子和生物大分子一样。目前绝大多数用于 mRNA 递送的非病毒载体是通过自组装制备的,其中包括很大一部分 mRNA 疫苗。
二、mRNA疫苗
mRNA疫苗可分为两种类型:
- 非复制型和
- 自我扩增(也称为自我复制和复制子)mRNA疫苗。
虽然这两种疫苗在 mRNA 结构中具有共同的结构,但自扩增 RNA 疫苗在RNA 复制的编码区包含额外的序列
2.1 . mRNA疫苗的优势
-
mRNA疫苗的第一个重要优势是安全性。与减毒活疫苗和灭活疫苗相比,mRNA 疫苗排除了与内毒素和感染相关的任何问题。与DNA 疫苗和基于病毒载体的疫苗不同,mRNA 疫苗不会带来基因组整合和插入诱变的风险,因为它们的活性不需要进入细胞核 [ 1 ]。mRNA 活性的瞬时性质有利于避免相应蛋白质的持续表达,从而更好地控制它们的活性。
-
mRNA疫苗的第二个好处是功效。由于 mRNA 疫苗提供必需的抗原,因此与全细胞疫苗相比,它们促进了对抗原更具特异性的适应性免疫的发展,并将不良反应降至最低。此外,mRNA 疫苗的药理学提供了细胞免疫反应和抗体反应的发展,这可用于治疗需要细胞介导免疫的疾病,例如癌症。
-
mRNA疫苗的第三个优势在于它们的生产。临床级 mRNA 的合成基于高度标准化的体外转录 (IVT) 过程。制造过程在无细胞系统中使用 DNA 模板和各种酶。与基于培养的 DNA 或蛋白质疫苗生产相比,该过程相对稳健且可扩展。此外,合成相似大小的新抗原序列所需的优化更少。mRNA 制造的这种灵活性对于生产针对快速传播的感染因子的 mRNA 疫苗非常有利。病毒刺突蛋白被释放 ,使I、II、III期人体临床试验以前所未有的速度启动:分别在病毒序列确定后的66天、140天和199天(NCT04283461、NCT04405076、NCT04470427)内,导致紧急情况一年内在美国获得其 mRNA 疫苗的使用授权 (EUA) [ 13 ]。
虽然前景广阔,但 mRNA 疫苗领域仍有改进的空间。mRNA等阴离子大分子由于来自细胞膜的静电排斥力而难以进入细胞,而mRNA由于细胞外空间中丰富的核糖核酸酶(RNases)容易降解[ 2 ]。此外,细胞外 mRNA 内源性地充当模式识别受体的配体,例如 toll 样受体 (TLR) [ 14 , 15 ] 和 RIG-I 样受体 (RLR) [ 16 , 17],这可能会造成不利影响。在这些方面,IVT mRNA 通常被包装在可以掩盖不良副作用并促进细胞进入的递送载体中,尽管有报道称裸 mRNA在体内[ 18、19 ] 和离体[ 20 ]有效。Geall 及其同事证明,将 mRNA 包装在基于脂质的纳米颗粒(LNP) 中可以保护 mRNA 货物免受 RNases [ 21 ]。他们还表明,封装在 LNP 中的 0.1 μg mRNA接种比接种 1.0 μg 裸 mRNA的体内转染增强 10 倍,这表明使用递送载体进行 mRNA 递送的好处 [ 21]。
2.2 . mRNA的结构元件
IVT mRNA的结构元件与天然mRNA相似,由5’帽、5’非翻译区(UTR)、编码区、3’UTR和poly(A)尾组成(图1A )。
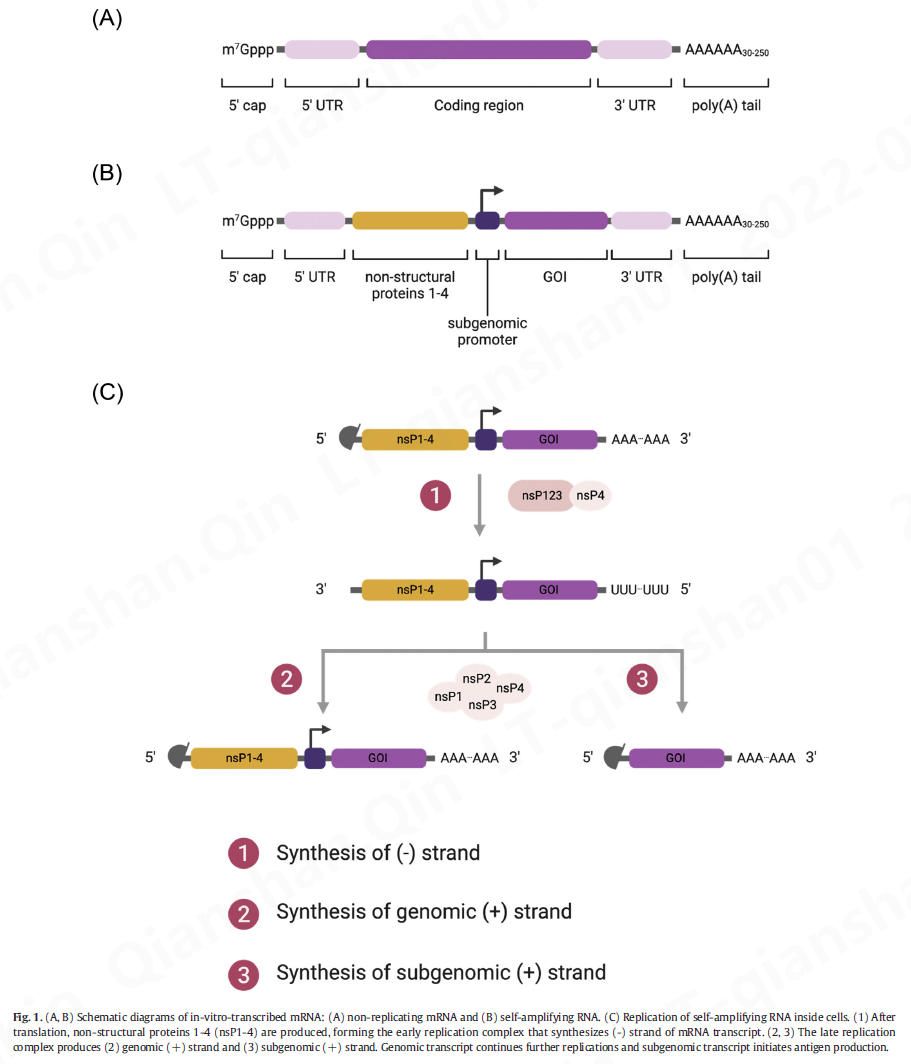
2.2.1 . 5' 帽
真核细胞质中的天然 mRNA 在 5' 末端含有 7-甲基鸟苷 (m 7 G) 部分,然后是与第一个核苷酸(m 7 GpppN)的三磷酸键。这种保护结构称为 5' 帽,它调节前 mRNA 剪接和核输出,保护 RNA 免受外切核酸酶切割,并启动mRNA 翻译[ 22 ]。5' 帽还参与先天免疫系统从自身 RNA 中识别非自身 RNA
。。。
2.2.2 5' 和 3' UTR
UTR 不翻译成蛋白质,但参与调节 mRNA 表达。5' 和 3' UTR 分别位于编码区的上游和下游。它们包含与 mRNA 的稳定性、核糖体对 mRNA 的识别、与翻译机制中的组件的相互作用以及 mRNA二级结构相关的各种调节序列[ 29 ]。在 mRNA 治疗中,在 UTR 中包含顺式调控序列可以改善 mRNA 的翻译和半衰期。已从天然存在的序列中鉴定出许多促进 mRNA 翻译的 UTR 序列。例如,源自 α 和 β 珠蛋白的 UTR 序列已广泛用于设计 IVT mRNA 构建体
2.2.3 . 聚(A)尾
mRNA 在其 3' 端有一个多聚腺苷酸化区域,称为多聚 (A) 尾。它是 mRNA 分子寿命的重要决定因素。哺乳动物细胞中天然 mRNA 分子的 Poly(A) 尾长度约为 250 个核苷酸 (nt),在胞质溶胶中 mRNA 的整个生命周期中,它们逐渐从 3' 缩短到 5' [ 37 ]。由于尾部大小通过调节 3' 核酸外切降解来影响 mRNA 的衰变,因此在 mRNA 治疗剂的生产中需要在大约 100 nt 处掺入 poly(A) 尾部。聚腺苷酸化主要有两种方法IVT mRNA:
- 使用重组多聚 (A) 聚合酶将酶促添加到加帽的 mRNA
- 嵌入质粒模板中的多聚 (T) 序列。
虽然poly(A)聚合酶介导的加尾适用于小规模生产,但这种方法受限于poly(A)尾长度不一致,并且在合成中增加了额外的步骤,不利于大规模生产。另一方面,在质粒模板中嵌入 poly(T) 序列可以在 IVT 反应过程中一步合成 poly(A) 尾,因为用于加尾的模板包含在质粒 DNA中. 此外,它会产生具有确定长度的 poly(A) 尾,这增加了 mRNA 治疗的可重复性。与此方法相关的一个困难是含有聚 (A:T) 区域的环状质粒往往难以克隆,并且在大肠杆菌中的 DNA 繁殖过程中易于缩短[ 38 ]。为了克服克隆的困难,Arbuthnot 等人。提出用 IIS 型酶重复限制性消化、连接和增殖,以在环状质粒中将同聚序列延长至 100 bp [ 39 ]。同样,格里尔等人。提出线性质粒作为mRNA 合成的模板,以规避限制 [ 40 ]。
2.2.4 . 修饰的核苷酸
天然 RNA 分子是基于四种基本核苷酸(ATP、CTP、GTP 和 UTP)合成的。在 mRNA 成熟过程中,mRNA 中的一些核苷酸被转录后修饰。这些天然存在的修饰核苷酸,如假尿苷和 5-甲基胞苷,可用于 IVT mRNA 的合成 [ 41 ]。使用这些修饰的核苷酸可以避免先天免疫系统识别 IVT mRNA,从而避免不良的免疫反应 [ 15 , 42 ]。Karikó 及其同事开创了用于产生 IVT mRNA 的修饰核苷酸,报告说在 IVT mRNA 中将尿苷替换为假尿苷不仅降低了针对 mRNA 的先天免疫反应,例如 TLRs。15 ] 和蛋白激酶 R [ 42 ],但也增加了蛋白质翻译[ 43 ]。5-甲基胞苷、2-硫尿苷和N 1 -甲基假尿苷被探索为碱性核苷酸的替代品,与未修饰的对应物相比,修饰的核苷酸的组合显示出更好的功效 [ [44]、[45]、[46] ]。然而,反例也有报道。塞斯等人。表明与假尿苷取代的 mRNA 相比,未修饰的 mRNA 在小鼠中表现出更高的蛋白质产生和温和的细胞因子诱导 [ 47]。考夫曼等人。证明当通过 LNP 静脉内给药时,与未修饰的 mRNA 相比, mRNA 的假尿苷取代不会改变体内蛋白质表达和各种细胞因子的水平 [ 48 ]。最近,Vaidyanathan 等人。报道了一项关于修饰核苷酸对Cas9 mRNA 活性、白细胞介素和 TNF-α 水平影响的比较研究[ 49 ]。他们没有发现修饰核苷酸的掺入与 Cas9 mRNA 活性之间存在相关性。尽管与未修饰的核苷酸相比,修饰的核苷酸适度降低了免疫反应,但细胞因子的水平受到 HPLC纯化的影响更大 . 总之,修饰核苷酸的效果似乎不一致,可能取决于实验设置,例如细胞类型、给药途径和递送载体。尽管如此,考虑到至少有 143 种天然核苷酸修饰,很难说修饰的核苷酸是调节细胞中 RNA 活性的重要决定因素 [ 50 ]。同时,更多的证据表明,RNA 修饰的异常存在或缺失可能导致人类疾病,这支持了对修饰核苷酸的益处进行全面评估的必要性
2.3 . 自扩增 RNA 疫苗
自扩增 RNA (saRNA) 疫苗利用源自RNA 病毒(主要是甲病毒)的 RNA 依赖性 RNA 聚合酶 (RDRP)来扩增递送的 RNA,从而增加抗原蛋白的产生。除了 mRNA 的正常成分外,saRNA 还包含编码 RDRP 成分的大开放阅读框(ORF),包括非结构蛋白 1-4 (nsP1-4) 和以下感兴趣的基因(抗原序列)亚基因组启动子(图 1 B)。nsP1、2、3 和 4 编码负责mRNA 加帽、NTPase/解旋酶/蛋白酶、宏结构域的蛋白质和 RDRP。Abu Bakar 和 Ng 最近的一篇评论总结了每个组件的详细信息
一旦 saRNA 被传递到胞质溶胶,它就会通过核糖体中的内源性翻译机制。nsP 系列的翻译导致前体多蛋白的产生,形成合成 RNA 负链的早期复制复合物。多蛋白的进一步切割产生晚期复制复合物,该复合物通过使用负链作为模板合成基因组和亚基因组 RNA 的正链。结果,一个 saRNA 拷贝产生多个 RNA 转录物拷贝(图1C ),克服了体内mRNA 治疗剂的有限细胞溶质递送。
saRNA 的抗原表达最初逐渐增加并持续较长时间 [ 53 ],从而延长了对抗原呈递细胞 (APC) 的刺激。
与 saRNA 疫苗相关的一个缺点是大而复杂的saRNA 序列。nsP1-4 序列的长度约为 7 kb,这通常使 saRNA 疫苗的全长超过 9 kb。最近减少 saRNA 构建体大小的努力包括将 nsP 系列的序列和感兴趣的基因拆分为单独的 RNA 分子,称为反式扩增或反式复制系统 [ 58]。由于其在多功能性、安全性和易于大规模生产方面的潜在优势,这种跨复制系统可能是有利的。贝塞特等人。比较了自扩增和反式扩增 RNA 疫苗的翻译效率,发现反式扩增 RNA 疫苗的 mRNA 翻译与 saRNA 疫苗一样有效,值得进一步探索反式扩增 RNA 疫苗 [ 59 ]。尽管反式扩增 RNA 系统的尺寸相对紧凑,但复制酶的 mRNA 长度仍然超过 7 kb,这需要具有大负载能力的传递载体才能成功传递 RNA。
三、mRNA疫苗的作用机制
mRNA 疫苗接种通过几种可能的途径导致获得性免疫:
- 转染体细胞,如肌肉细胞和表皮细胞,
- 转染注射部位的组织驻留免疫细胞
- 转染免疫细胞在次级淋巴组织中,包括淋巴结(LNs)和脾脏(图 2 )。
通过亲代途径施用的mRNA 疫苗,例如皮内、肌肉内和皮下注射,可以转染注射部位附近的非免疫细胞 [ 60 , 61 ]。非免疫细胞转染导致抗原的产生,随后在蛋白酶体中降解. 降解后,源自抗原的表位可与I 类主要组织相容性复合物(MHC) 形成复合物,用于将抗原呈递给表达 CD8的细胞毒性T 细胞,从而建立对抗原的细胞免疫。还已知肌细胞的转染会激活骨髓来源的 DC,然后是 CD8 + T 细胞引发 [ 62 ]。虽然确切的机制尚不清楚,但认为抗原从肌细胞转移到 DC

mRNA 疫苗还可以转染组织驻留的免疫细胞,主要是APC,例如 DC 和巨噬细胞 [ 63 ]。mRNA 疫苗的肠胃外给药可以在注射部位引发局部免疫反应 [ 2 , 64 ]。这会将免疫细胞募集到该区域,促进组织驻留免疫细胞的转染,例如 DCs 和巨噬细胞 [ 63 , 65 ]。与第一种情况一样,将 mRNA 转染到这些细胞会导致抗原呈递在 MHC I 类上,从而导致 CD8 + T 细胞成熟(图 3)[ 2 , 66]。此外,APCs 可以通过 MHC II 类途径处理抗原,因此 APCs 的 mRNA 转染可以导致表达 CD4的T 辅助细胞的激活(图 3)[ 2 ]。否则,mRNA 疫苗会通过淋巴引流并通过淋巴系统转运到邻近的 LN [ 63、66 ] 。LN 是各种免疫细胞(包括单核细胞和幼稚 T 细胞和 B 细胞)驻留的部位,位于这些次级淋巴器官中的抗原会引发适应性免疫反应。在 LN 中,mRNA 疫苗转染 LN 驻留细胞,例如 APC 和内皮细胞[ 63、66、67]。这些细胞的转染不仅可以启动 T 细胞,还可以启动 B 细胞。
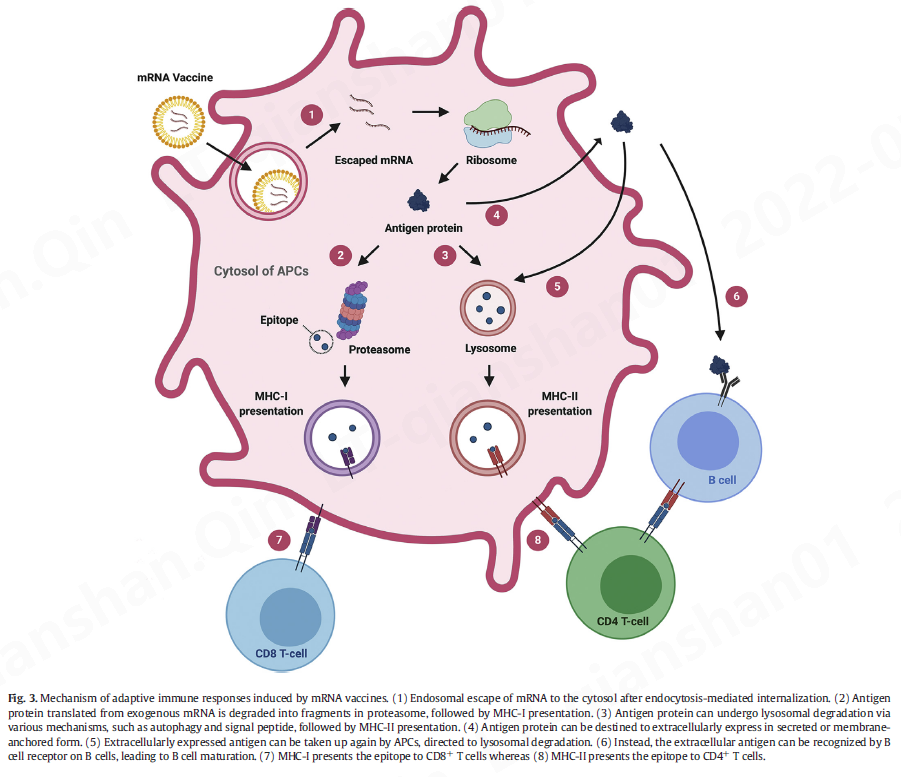
为了引发针对抗原的适应性免疫,mRNA 疫苗必须通过内源性翻译机制翻译成抗原(图 3)。由于翻译后的抗原最初位于胞质溶胶中,因此细胞优先将其视为内源性抗原。这意味着由 mRNA 疫苗产生的内源性抗原很容易呈递在 MHC I 类分子上,从而引发 CD8 + T 细胞 [ 2 ]。 然而,启动 CD4 + T 细胞对于建立有效和强大的细胞免疫也是必不可少的 [ 68 ]。
此外,考虑到 T 辅助细胞在 B 细胞启动中的作用很重要 [ 69 ],mRNA 疫苗启动 CD4 +T 细胞对于发展体液免疫也是非常可取的。如前所述,mRNA 疫苗接种导致 CD8 + T 细胞和 CD4 + T 细胞的激活,这表明转染的 APC 具有将内源性抗原加工成 MHC II 类呈递的替代途径 [ 68 ]。在 MHC II 类上呈递内源性抗原的一个例子是通过与细胞溶质抗原的溶酶体降解相关的自噬 [ 70 ]。众所周知,内源性抗原在 MHC II 类上的呈递可以由热休克蛋白 90和与抗原加工(TAP) 复合物相关的转运蛋白作为非自噬途径介导。71 ]。MHC II 类上的内源性抗原的抗原呈递可以通过工程化mRNA 序列进一步增强(图 3 )。通过编码将蛋白质转运到特定细胞内区室的信号肽,可以增强 MHC II 类上的抗原呈递 [ 72 ]。例如,Bonehill 等人。在抗原序列之前插入不变链的靶向序列,以编程要在 MHC II 类上呈递的翻译蛋白质 [ 73 , 74 ]。其他信号肽,如 MHC I 类 [ 75 , 76 ] 和 II [ 77 , 78 ]] 和溶酶体相关膜蛋白-1 (CD107a) [ 68 , 79 ] 可以掺入 mRNA 编码区以引发抗原特异性 CD4 +和 CD8 + T 细胞。或者,可以设计mRNA序列以在细胞外表达抗原(图3 ),以分泌形式[ 11、80、81 ]或跨膜形式[ 11、81、82 ]。例如,Richner 等人。通过编码病毒的 ME 蛋白研究了针对寨卡病毒的mRNA 疫苗,该蛋白在小鼠中产生病毒样亚病毒颗粒 [ 80]。蒂瓦里等人。工程化的 mRNA 编码来自衰变加速因子的糖基磷脂酰肌醇 膜锚定序列,以表达锚定在质膜上的感兴趣的蛋白质 [ 83 ]。科比特等人。设计了 mRNA 序列,将 SARS-CoV-2 的融合前稳定的刺突蛋白编码为跨膜锚定蛋白 [ 11 ]。他们还比较了两种 mRNA 疫苗对 MERS 的功效——一种编码分泌形式的 MERS 刺突蛋白,另一种编码跨膜锚定的 MERS 刺突蛋白——并表明后者引发了更有效的假病毒中和抗体反应. 尽管如此,很难概括跨膜形式比分泌形式更有效。因此,用于抗原表达的 mRNA 构建体——无论是分泌形式还是膜锚定形式——都可以针对单个病毒株进行定制,以最大限度地提高疫苗的安全性和有效性。细胞外表达的抗原可以被 APC 识别,并分别通过 MHC II 类呈递和交叉呈递有效地引发 CD4 +和 CD8 + T 细胞反应。值得注意的是,mRNA 衍生的细胞外抗原也有助于产生抗原特异性体液反应 [ 84 , 85 ]。
抗体反应是疫苗中和外来抗原的关键机制。在 mRNA 疫苗的情况下,B 细胞的适当激活仍然是诱导和维持抗体产生的关键。B细胞活化是通过在细胞外空间与B 细胞受体(BCR) 相遇的完整抗原引发的(图 3 )。由于幼稚 B 细胞存在于 LN 中,因此 B 细胞激活需要将抗原转运到 LN (Lymph node)以进行相遇。可溶性抗原从外周到达引流淋巴结;然而,小蛋白质(尺寸小于 10 nm)往往会进入毛细血管,因为血液流速比淋巴流速高 100-500 倍率,限制了 LN 中的抗原可用性 [ 86 ]。单核细胞和迁移性 DC 等 APC 将抗原携带至 LN 并将其提供给幼稚 B 细胞;然而,与将抗原直接递送到 LN 相比,它的效率相对较低 [ 87 ]。因此,可以靶向 LN 内细胞的 mRNA 疫苗可以通过增加 LN 中抗原的局部浓度来改善抗体反应 [ 87 ]。虽然 LN 中的可溶性抗原可以被 B 细胞直接识别,但它们可以被其他细胞捕获以激活 B 细胞,从而使 B 细胞更有效地识别抗原 [ 88]。根据抗原是否被调理,包膜下窦巨噬细胞或 DC 可以捕获抗原并呈现在其膜上以激活 B 细胞 [ 87 , 89 ]。
BCR 结合抗原通过内吞作用被内化、消化并呈递在 MHC II 类上。之后,B 细胞可以分化为短寿命浆细胞,这些浆细胞迅速分泌低亲和力抗体或进入生发中心 (GC),在那里它们发生体细胞超突变和亲和力成熟。进入 GC 的活化 B 细胞将 MHC II 类抗原呈递给 T 辅助细胞,并在 T 细胞/B 细胞边界处接收共刺激信号。随后,它们在 GC 的暗区经历体细胞超突变,增殖并磨练 BCR 对抗原的特异性。然后,GC B 细胞离开到光区,在那里它们通过与滤泡 DC 的相互作用进行亲和力成熟。接受来自 B 细胞和 DC 的抗原的滤泡 DC 将抗原储存在它们的非降解隔室中,并将抗原长期呈递给 B 细胞,帮助它们的亲和力成熟。亲和力成熟的循环级联能够选择具有高亲和力 BCR 的 B 细胞,并通过细胞凋亡清除具有低亲和力 BCR 的 B 细胞。记忆 B 细胞[ 69 , 90 ]。通过疫苗接种建立这两种细胞是产生高亲和力抗体和确保针对抗原的长期记忆的关键。在二次暴露于特定抗原后,记忆 B 细胞会重新激活抗体产生,从而实现快速的抗体介导的免疫反应 [ 91 ]。主要存在于骨髓中的长寿浆细胞提供长效的保护性体液免疫,持续数年甚至数十年[ [91]、 [92]、 [93] ]。Laczkó 等人。最近证明,单剂量针对 SARS-CoV-2 的 mRNA 疫苗产生抗原特异性记忆 B 细胞和长寿浆细胞,支持 mRNA 疫苗产生持久体液免疫的能力 [ 94 ]。
因此,理想的 mRNA 疫苗不仅可以激活 CD8 +和 CD4 + T 细胞,还可以为 LN 提供抗原以激活 B 细胞并产生针对病原体的抗体反应。通过编码信号肽或细胞外形式的抗原,可以对 mRNA 构建体进行工程改造以引发更有效的 T 细胞启动。此外,考虑到 GC 反应在抗体反应中的重要性,设计有效 mRNA 疫苗的一种方法可能是用有效的递送载体靶向 LN [ 95 ]。
四、给药途径
疫苗接种的给药途径会对mRNA 疫苗的功效产生影响。疫苗给药的方式会影响免疫细胞对引发的可及性以及疫苗接种后后果效应(局部或全身免疫反应)的程度。尽管给药途径有明显影响,但对给药途径对 mRNA 疫苗时空分布影响的基本理解仍然有限。
4.1 . 肌肉内、皮下和皮内给药
肠胃外给药(Parenteral administration)是疫苗接种的普遍途径。大多数疫苗通过侵入性程序施用,例如肌肉内 (IM)、皮下 (SC) 和皮内 (ID) 注射 [ 96 ]。这些给药途径需要较少的培训来执行,并且被认为比静脉 (IV) 途径更安全地避免潜在的感染风险 [ 97 ]。由于存在大量APC ,如朗格汉斯细胞,皮肤和肌肉组织是很有吸引力的疫苗接种部位。此外,对这些组织的给药使疫苗能够进入淋巴管,从而增强疫苗向 LN 的递送。因此,通过 IM、SC 和 ID 注射施用的 mRNA 疫苗可以通过转染组织驻留细胞和 LN 驻留细胞来引发强烈的免疫反应。最近,Santangelo 及其同事使用 PET-CT 和红外成像观察了在IM 注射到食蟹猴后,用双放射性核素-NIR 探针标记的 mRNA 疫苗的早期运输[ 63]。mRNA 疫苗到达从注射部位排出的 LN;然而,它们很少到达远端 LN。这一结果证实了之前关于蛋白质疫苗的报道 [98,99 ]。此外,主要在单核细胞中观察到 mRNA 转染,在注射的肌肉组织和引流 LN中的 DC、B 细胞和 CD4 + T 细胞中观察到的程度较小,这与之前的发现一致 [ 65 ]。这表明 mRNA 疫苗接种可能涉及单核细胞转染作为主要机制,尽管这可能是由于研究中使用的mRNA 载体的特性。尽管SC 注射被认为比 IM 注射更具免疫原性,因为注射部位靠近含有大量组织驻留免疫细胞的真皮 [ 100 ],对基于蛋白质的疫苗进行的多项独立研究表明,肌肉注射和皮下注射之间的抗体滴度没有显着差异 [ 99 , 101 ]。尽管如此,接种疫苗的注射方法被证明会影响疫苗在非人类灵长类动物中的生物分布[ 99 ] 和 GC 反应 [ 101 ]。与基于蛋白质的疫苗的结果一致,在小鼠中与 mRNA 疫苗进行的比较研究表明,肌肉注射和皮下注射之间的抗体产生没有显着差异 [ 102]。
由于真皮中有大量免疫细胞,ID 给药被认为是一种有前途的疫苗接种途径;然而,由于给药技术困难,它的使用频率低于 SC 和 IM 注射。 然而,很明显,通过 ID 注射施用的 mRNA 疫苗可以产生更强烈的体液反应[ 102 ]。Loré 及其同事表明,ID 注射封装在 LNP 中的 mRNA 疫苗导致B 细胞反应在非人类灵长类动物中快速和扩展表达,这表明 ID 注射 mRNA 疫苗可能有利于提供对流感病毒的适应性免疫的快速发展[ 65、84 ]。_
4.2 . 静脉给药
静脉内 (IV) 注射 mRNA 治疗剂可导致体内编码蛋白质的最高产量。通常,静脉注射的 mRNA 治疗剂靶向肝脏,有效地转染肝细胞、枯否细胞和内皮细胞 [ 60 ]。然而,在疫苗接种中,IV 注射不太受欢迎,因为 mRNA 疫苗在循环系统中的广泛生物分布,可能导致全身副作用。尽管如此,通过将脾脏(另一个次级淋巴器官)作为转染位点,从 IV 注射的 mRNA 疫苗中产生适应性免疫反应是可以实现的 [ 103 , 104 ]。克兰兹等人。证明含有_癌症抗原mRNA 在 IV 注射后转染脾脏,导致小鼠产生抗肿瘤免疫 [ 105 ]。值得注意的是,他们表明 mRNA 脂质复合物的表面电荷改变了从肝脏、脾脏到肺部的生物分布和转染。此外,他们证明,与 SC 注射相比,静脉注射 mRNA-lipoplex 疫苗可引发更强大的 CD8 + T 细胞启动。为了治疗黑色素瘤和三阴性乳腺癌(NCT02410733 和 NCT02316457),这种全身性 mRNA 方法的临床评估正在进行中。尽管取得了令人鼓舞的结果,但 mRNA 疫苗的 IV 给药仍需要进一步优化,以增强靶特异性 mRNA 递送并最大限度地减少全身副作用 [ 106]。
或者,可以利用静脉注射 mRNA 治疗剂从肝脏产生抗体和蛋白质,以中和目标病原体或癌细胞 [ [107]、[108]、[109] ]。这种策略源于基于 mRNA 的蛋白质替代疗法的概念,该疗法使用肝脏作为生物工厂来生产免疫蛋白。Rybakova 等人。改造mRNA 序列以编码曲妥珠单抗的重链和轻链,曲妥珠单抗是一种靶向癌细胞上 HER2 受体的单克隆抗体,并证明静脉注射 mRNA-LNP 可导致小鼠肝脏体内产生曲妥珠单抗 [ 108]。我们还报道了 mRNA-LNP 可用于在肝脏中产生可溶性血管紧张素转换酶 2 (ACE2),通过与刺突蛋白结合来中和 SARS-CoV-2 病毒 [ 109 ]。这些结果表明,全身给药mRNA 疗法提供了一种基于单克隆抗体的免疫疗法的替代方法。
4.3 . 鼻内给药
另一种疫苗接种方法是粘膜疫苗递送,靶向非胃肠道粘膜淋巴组织和淋巴结。施用到黏膜层(例如鼻和肺黏膜)的 mRNA 疫苗通过上皮下的淋巴系统进入引流黏膜 LN [ 86 , 110 , 111 ]。特别是,预防性 mRNA 治疗以预防呼吸道疾病,如流感 [ 112 ] 和RSV [ 83 ],优选通过鼻内 (IN) 给药 [ 113 ]] 因为粘膜疫苗接种可以提供病原体特异性抗体,这些抗体可以分泌到粘液中,这些抗体可以在感染的早期阶段中和病原体 [ 114 ]。此外,mRNA 疫苗的粘膜递送可导致免疫球蛋白 A (IgA) 的分泌。IgA 通常以二聚体形式存在于粘液中,通过阻断和中和细菌毒素和病毒,在抵抗感染的前线发挥作用[ 115 ]。例如,李等人。据报道,IN 给药的基于 PEI 的聚合物纳米颗粒将 HIV gp120 mRNA 传递到鼻上皮,导致小鼠体内产生抗原特异性 IgG 和 IgA [ 113 ]。
IN 给药要考虑的一个方面是传递载体的粘液渗透。粘液通过去除异物来保护上皮细胞,这也有助于去除鼻内给药的纳米颗粒。鉴于粘弹性和粘性粘液可以阻止 mRNA 疫苗转染细胞,开发粘液穿透载体可能有利于 IN 给药后有效的 mRNA 疫苗接种。众所周知,优化纳米粒子的物理化学性质可提高粘液渗透性 [ 116 , 117 ]。制造纳米颗粒粘液穿透或粘液惰性的策略在其他地方进行了详细审查 [ 116 , 117]。最后,在 IN 给药后对 mRNA 疫苗的大多数评估仅限于啮齿动物模型,敦促进一步研究 IN 给药在较大动物中的优势。
综上所述,mRNA疫苗给药途径的选择会影响mRNA疫苗的安全性和有效性。因此,考虑到疾病及其发病机制,可以确定 mRNA 疫苗接种的适当给药途径。此外,了解每条递送途径的生物学特性将促进 mRNA 疫苗递送载体的合理设计。
五、 mRNA的细胞内递送
对于有效的疫苗接种,mRNA 疫苗必须到达可以产生抗原的细胞质,从而触发针对抗原的适应性免疫。然而,如前所述,mRNA 分子是一个大的聚阴离子,因为其骨架中存在磷酸基团,阻碍了其直接进入细胞质。出于这个原因,纳米级生物材料因其能够以安全有效的方式将 mRNA 有效载荷偷运到细胞中而对 mRNA 疫苗的递送产生了极大的兴趣。
纳米级 mRNA 载体的细胞进入主要由内吞作用介导,内吞作用是细胞运输途径的相互连接的网络。纳米载体的内吞作用在纳米医学领域被广泛讨论[ [118]、 [119]、 [120]、 [121] ]。胞吞作用是由质膜和靠近细胞的纳米载体之间的动态非共价相互作用引发的。众所周知,纳米载体的物理化学性质,如尺寸、形状、表面电荷和表面组成,会影响确定的内吞作用途径——巨胞饮作用、小窝蛋白介导、网格蛋白介导、网格蛋白和小窝蛋白独立 [ 122 , 123 ]。内吞作用后,内化的纳米载体被转运到早期内体,然后成熟为多泡晚期内体。晚期内体进一步发展为溶酶体,其中大多数纳米载体被消化酶降解。否则,内化的纳米载体可以在内溶酶体运输过程中通过回收途径消除。大约 70% 的封装siRNA的内化LNP通过回收途径通过胞吐作用被清除,从而限制了遗传货物的细胞溶质传递 [ 124]。虽然 mRNA-LNP 的回收利用可能基于它们的大小和组成相似,但迄今为止尚未报道载有 mRNA 的 LNP 的内吞回收。从回收和降解中仅回收少量内化的纳米载体,并最终到达胞质溶胶。从数值上看,Gilleron 等人。证明当它们驻留在早期和晚期内体中时,只有 2% 的 siRNA 负载 MC3 LNP 在有限的时间窗口内逃逸到细胞质 [ 125 ]。最近,Sabnis 等人。据报道,包裹 mRNA 的基于 MC3 和脂质 5 的 LNP 分别表现出 2.5% 和 15% 的内体逃逸效率 [ 126 ]。尽管无数次努力破译内体逃逸的机制,但对机制的理解仍然有限。
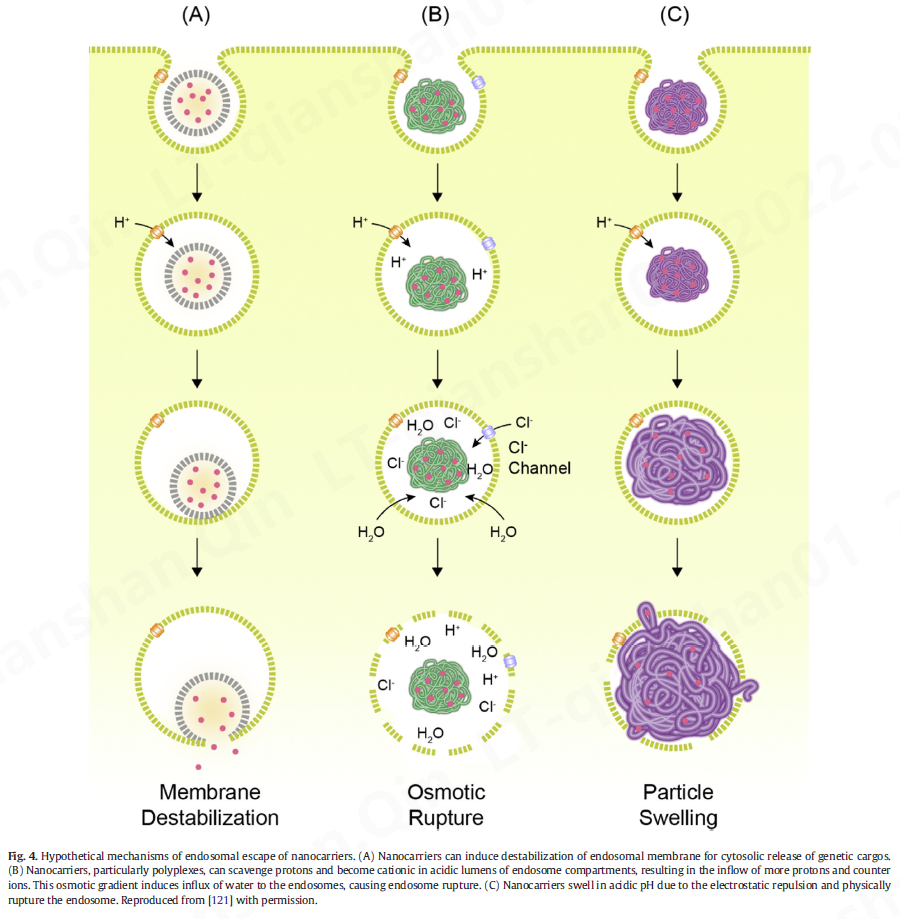
内溶酶体途径最显着的特征是pH的梯度变化。出于这个原因,促进纳米载体内体逃逸的主要方法是使它们对内体的酸化有反应。在诱导纳米载体内体逃逸的各种策略中,三种情况被广泛接受:(1)内体膜不稳定,(2)“质子海绵”效应导致内体渗透性破裂,以及(3)内体破裂通过颗粒膨胀(图 4)。一般而言,基于脂质的材料被认为具有更高的破坏膜稳定性的倾向,并且聚合物材料被认为通过渗透或肿胀介导的破裂来破坏内体[ 121]。无论材料和机制如何,内体逃逸的效率仍然极低,这需要实施新的方法来捕捉内体逃逸的复杂性。
六、 用于 mRNA 递送的材料
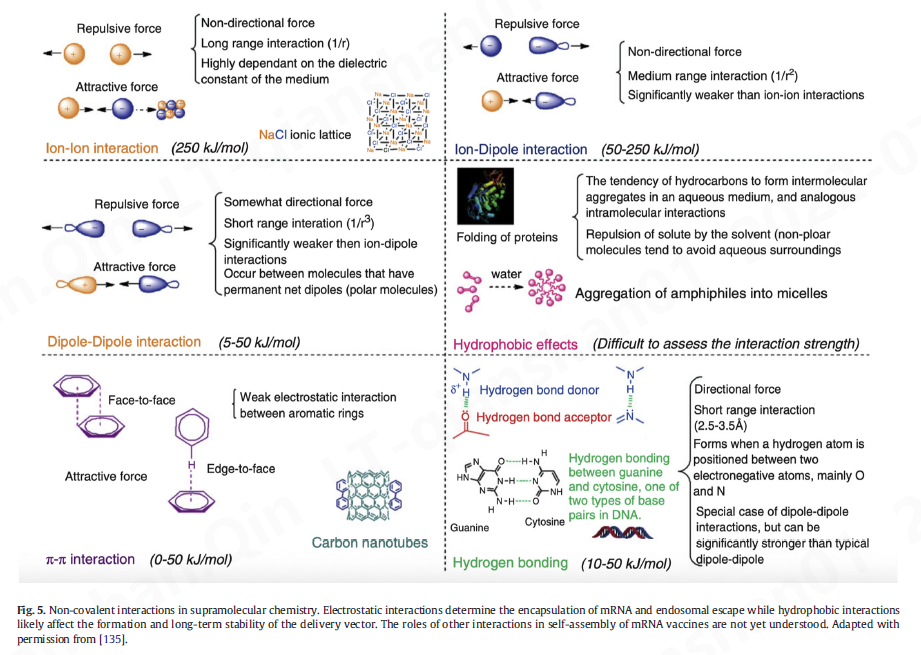
“智能”材料的自组装在材料科学的许多领域中是一种备受追捧的方法——只需混合组件,分子间的相互作用就会将这些组件组装成具有所需特性的所需结构。这种自发组织通常受非共价分子间相互作用的控制,例如静电、疏水、范德华和 π 效应(图 5)[ [133]、[134]、[135] ]。自组装的使用已在纳米科学的众多领域 [ 7 ] 中得到应用,包括纳米医学。在基因传递中,小分子等构建模块或聚合物与核酸相互作用并自组装成有序结构。
由于其多功能性,自组装在 mRNA 递送载体的设计中主要受到青睐。然而,自组装的主要挑战是确定构建块的必要特性,这些特性将转化为自组装基因传递载体的所需特性。这些特性是:
- 保护核酸免受核酸酶的影响,
- 核酸的受控释放
- 细胞和组织选择性,
- 高递送率
- 最小毒性
- 稳定性,尤其是在长期储存中。
尽管在基因传递载体的设计上付出了巨大的努力,但构建块特性对有效细胞摄取的独立影响仍不清楚 [ 136 ]。
迄今为止,载有 mRNA 的自组装纳米结构比裸 mRNA 疫苗产生更高的递送效率,并允许更广泛的给药途径,扩大了 mRNA 纳米药物的治疗潜力 [ 137 ]
6.1 . 基于脂质的系统
虽然问题仍然是必须精确定制哪些参数才能最大限度地传递特定核酸有效载荷,但很明显,LNP 的结构因核酸有效载荷 [ 147 , 151 , 152 ] 并由自组装的初始阶段而异。
LNPs 的自组装是由动力学和热力学之间持续的竞争驱动的,使它们成为高度流动和动态的系统。从历史上看,脂质囊泡通常通过脂质薄膜的再水化和随后的挤压来制备,以实现均匀的尺寸分布 [ [155]、[156]、[157]、[158] ];在这种情况下,脂质囊泡的形成可能是由热力学驱动的,因为脂质在水性介质中的溶解度很差。结果,这些脂质囊泡对核酸货物的封装较差[ 159 ]。如今,LNPs 是通过快速混合制备的,通常由微流控装置促进[153、154、160、161 ]。_ _ _ _ _ 一种特别流行的微流体制备方法是乙醇稀释,指的是当将乙醇溶液添加到过量的水性介质中时,脂质会迅速凝结成纳米液滴[ 159 ]。得到的 LNP 可以被认为是一种动力学产物,通常会产生大量的核酸包封。
通常,LNP 被认为是通过合并较小的脂质囊泡 [ 153 ] 的级联形成的,类似于经典的 LaMer 模型(成核 - 生长 - 成熟)。LNP 种子的初始形成是由阴离子核酸和带正电荷的脂质之间的静电相互作用指导的。对于负载 siRNA 的 LNP,建议种子具有类脂质体结构,其中 siRNA 夹在带电的可电离脂质层之间,生长导致中性可电离脂质填充纳米颗粒的核心 [ 153]。库尔卡尼等人。在一项使用基于 FRET 的融合测定的机制研究中表明,LNP 的生长更多地依赖于 pH 中和以及中性、未结合的可电离脂质向 LNP 核心的迁移,而与有效载荷(mRNA、微型环状 DNA 或 pDNA)无关 [ 162 ]。根据他们的研究,生成的 LNP 的内部结构通常可能是层状的,甚至可能是“洋葱状”。事实上,已经观察到 mRNA-LNP 的洋葱状结构,尽管没有迹象表明 mRNA 在脂质层之间被捕获 [ [163]、[164]、[165] ]。另一方面,mRNA-LNPs 可能形成倒六角形内部结构 [ 166]。Yanez Arteta 等人。表明可电离脂质、胆固醇和萤火虫荧光素酶mRNA 的混合物根据溶剂环境形成不同的结构,在类似于传统乙醇稀释方法的条件下组装成倒置的脂质小管 [ 166]。这一观察结果表明,LNP 的生长可能从反向胶束状种子结构开始,其余的脂质聚集到纯粹基于疏水相互作用的那些。无论种子结构如何,疏水相互作用似乎是 LNP 生长的主要驱动力,而静电相互作用指导种子形成和最终组装的稳定性。种子和最终结构的差异可能来自有效载荷的大小(例如,siRNA 是 20-30个核苷酸,而 mRNA 是大约 1000 个或更多核苷酸)或它们的灵活性(siRNA 是刚性双链体,而 mRNA 是柔性单链体)链)。此外,各种mRNA 序列可能形成局部二级结构取决于核苷酸序列 [ 167,168],这可能会干扰 RNA-脂质复合。迄今为止,核酸有效负载的性质对 LNP 自组装的影响以及相应优化脂质制剂以适应 mRNA 的因素仍然难以捉摸,需要进一步研究。
有趣的是,核酸不必嵌入 LNP 的核心以防止核酸酶降解 [ 169 ]。此前,布里托等人。据报道,当将 saRNA 添加到预制纳米乳液中时,阳离子纳米乳液可以保护 saRNA 而不会失去功能 [ 55 ]。这些纳米乳液的冷冻电镜显微照片表明形成了各种大小的均质结构,这可能表明 RNA 位于液滴内部或包裹在带电表面周围。这一发现可能表明任何导致 mRNA脂质复合物形成的自组装机制都可以防止RNase降解,可能是通过选择性地干扰 RNase 识别机制而不影响翻译。
LNP 可以说是迄今为止最复杂的 mRNA 递送,其结构和组成脂质的数量高度可变。LNP(用于 siRNA 递送)的当前临床标准——第一个临床批准的含有 DLin-MC3-DMA、DSPC、胆固醇和 DMG-PEG-2000 的制剂 [ 138 ]——每个颗粒包含近 60,000 个脂质分子(图 6一个)。配方中的每种脂质都有不同的功能,这将在下面讨论。脂质的化学结构定义了它们的分子几何形状,这对于脂质包装、纳米结构的形成至关重要, 和生物活性。由于用于 mRNA 递送的基于脂质的材料在过去几十年中迅速发展,其结构的复杂性以及对疫苗制造的合成限制的考虑值得详细讨论。
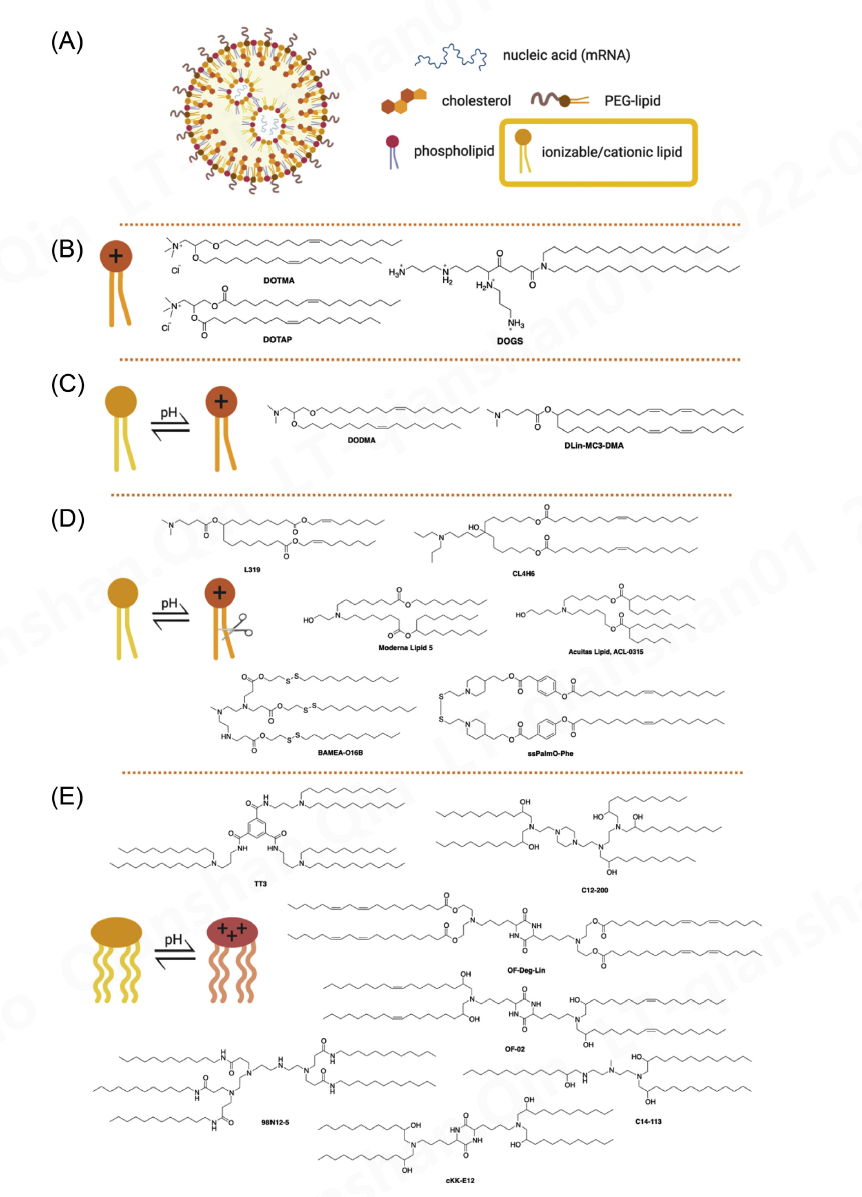
Fig. 6. (A) General structure of lipid nanoparticles. Ionizable or cationic lipids (yellow box) are the main component responsible for the encapsulation of nucleic acid and intracellular delivery. These lipids may be divided into groups shown below. (B) Structures of first-generation cationic lipids DOTMA, DOTAP, and DOGS; (C) Structures of ionizable lipids DODMA and DLin-MC3-DMA; (D) Structures of selected ester-based and disulfide-based biodegradable ionizable lipids; (E) Structures of selected ionizable lipidoids.
阳离子脂质是为 mRNA 递送而开发的第一代脂质。它们含有一个季氮原子,使它们带有永久正电荷,或者含有一个伯胺,在生理 pH 值或低于生理 pH 值时赋予正电荷。正电荷使这些脂质与带负电荷的 mRNA 形成离子相互作用,形成脂质复合物,通常称为 lipoplex [ 170 ]。阳离子脂质的例子包括 DOTMA [ 171 ]、DOTAP [ 172 ] 和 DOGS [ 173 ](图 6 B)。基因递送的早期尝试使用完全由阳离子脂质组成的制剂。虽然早期的阳离子脂质在体外显示出有希望的基因传递,由于它们的永久电荷和不可生物降解的特性,它们可能会出现有限的体内功效和细胞毒性问题 [ 3 ]。通过掺入结构脂质(例如 DOPE)可以降低细胞毒性。Lipofectin是 DOTMA 和 DOPE 的混合物,是第一个在mRNA的体内翻译中被证明成功的 LNP 制剂之一 [ 60 ]。克兰兹等人。使用 Lipofectin RNA-lipoplex 靶向 DCs 用于小鼠的癌症免疫治疗[ 105 ],以及用于治疗晚期黑色素瘤的人类(NCT02410733)。
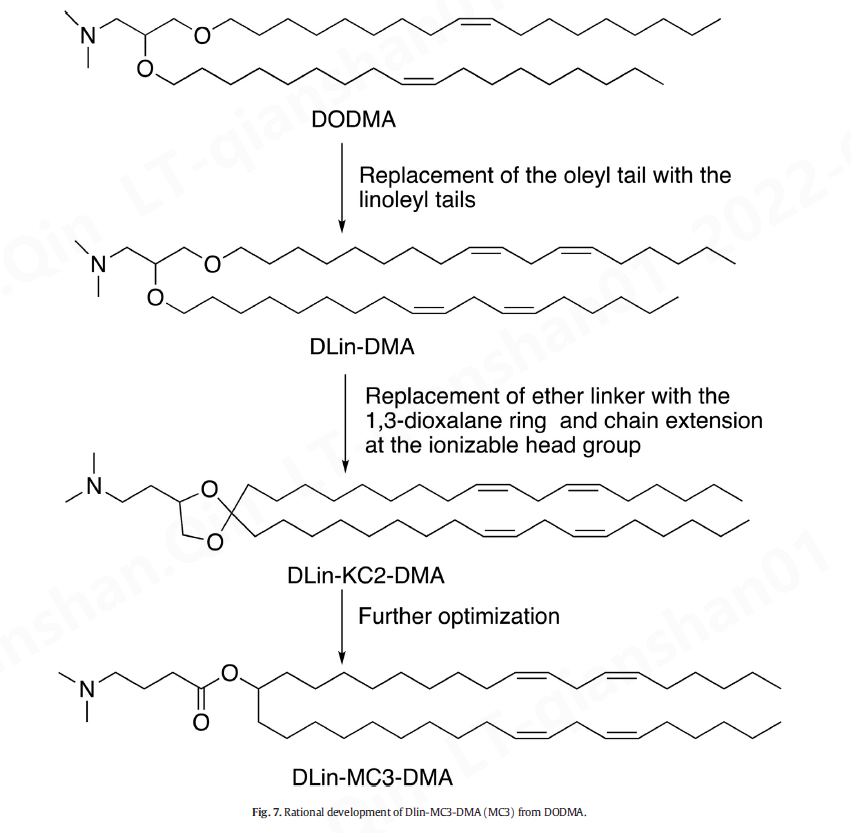
可电离脂质 (IL) 是第二代阳离子脂质,也称为 pH 依赖性脂质,在生理 pH 值下几乎没有电荷,但在酸性 pH 值下带正电荷。IL 的 pH 敏感特性可防止其通过网状内皮系统 (RES) 进行代谢,从而延长其半衰期 [ 174 ]。IL 也不太可能引起免疫激活或与血清蛋白的相互作用,从而提高其安全性 [ 3 ]。DODMA 是首批评估基因传递的 IL 之一(图 6 C 和7)[ 175 ]。用亚油基尾取代 DODMA 的油基尾导致 DLinDMA [ 175 , 176] 它在体内引发了针对呼吸道合胞病毒(RSV)的广泛、有效和保护性免疫[ 21 ]。DLinDMA 结构上的系统 SAR 导致识别 DLin-KC2-DMA,进一步优化导致发现 DLin-MC3-DMA,这是第一个 siRNA 药物 Onpattro® 的关键成分(图6C和7) [ [177]、[178]、[179]、[180] ]。因此,MC3 被证明在传递 mRNA 方面也很有效 [ 166、[181]、[182]、[183] 、[184]、[185]、[186]、[187]、[188] ]。MC3 具有 72 小时的长血浆半衰期,这限制了其长期给药 [ 126 , 189 ]。
因此,下一代脂质采用可促进清除的可生物降解官能团。增加生物降解性的一种方法是包含酯部分。酯基序易于结合、化学稳定、可生物裂解,并且可以有效控制其生物降解动力学[ [189]、[190]、[191] ]。基于酯的可生物降解脂质的一些例子包括 L319 [ 179 , 189]、YSK12-C4 [ 192 ]、CL4H6 [ 193 ]、Moderna 专有脂质系列 [ 126 , 194 ] 和 Acuitas 专有脂质系列 [ 195 ](图 6 D)。与 MC3 相比,这些可生物降解的酯基 ILs 经常被发现在基因传递中更有效。例如,与小鼠中的 MC3 相比,L319 表现出更好的基因沉默 [ 189 ]。同样,发现 Moderna 脂质 5 的效力是 3 倍 [ 126 ] 和 Acuitas 脂质 ACL-0315(用于辉瑞/BioNTech COVID-19 疫苗的 IL)在递送萤火虫荧光素酶方面的效力大约是 MC3 的 6 倍动物的 mRNA [ 195]。引入生物降解性的另一种方法是将二硫键添加到主链中。二硫键被谷胱甘肽(GSH) 或细胞中存在的其他二硫键还原酶生物还原 [ 196 ]。含有生物可还原二硫键的 IL 的实例包括 BAMEA-O16B [ 197 ] 和 ssPalmO-Phe 脂质(图 6 D)[ 198 ]。
衍生自可电离脂质和树枝状聚合物的混合分子称为类脂质。这些第三代阳离子脂质通常通过简单的化学方法合成,涉及的步骤数量有限,因此能够实现高通量转变 [ 199 ]。阿金克等人。报道了第一个类脂质 98N12-5 [ 200 ] 的例子,该例子经过进一步修饰,导致发现了优越的类似物 C12-200 [ 201 ] 和 C14-113(显示出对心脏的选择性)[ 201 , 202 ]。在随后的研究中,Dong 等人。发现 cKK-E12 作为一种有效的 siRNA 递送剂 [ 203]。cKK-E12 LNP 已被用于各种治疗目的,例如癌症免疫治疗 [ 66 , 108 ] 和基因组编辑 [ 204 ]。cKK-E12 脂质尾部周围的修饰导致脂质 OF-02 [ 205 ] 和 OF-Deg-Lin。后者对脾脏具有惊人的选择性(约 85%)并且能够转染 B 细胞,突出表明小的结构变化可以导致组织选择性 [ 206 ]。在另一项研究中,Li 等人。报道了 TT3 作为编码因子 IX [ 207 ]、CRISPR/Cas9 [ 208 ] 以及最近针对 SARS-CoV-2 的病毒抗原的各种 mRNA 分子的有效递送剂[34 ]。类脂质的结构列于图 6 E。
LNPs 通常还包含结构脂质(通常是两种——一种磷脂和胆固醇)和一种 PEG-脂质(图 6 A)。这些脂质的主要目的是提供颗粒稳定性、尺寸控制和血液相容性 [ 209 ];然而,它们在配方和化学性质中的数量可能需要调整,以从标准 MC3 配方中有效地传递 mRNA [ 207 , 210 ]。根据给药途径,也可能需要进行调整。例如,我们研究了是否用天然存在的甘油衍生物代替磷脂 DSPC可以改变 mRNA 递送效率,并发现引入 DGTS 代替 DSPC 可改善通过 IV 注射向肝脏的递送,但会降低雾化 LNP 对 IN 递送 mRNA 的功效 [ 188 ]。虽然尚不清楚如何利用这些变化的力量,结构性脂质可能位于 mRNA-LNP 的外围 [ 166 ],众所周知,脂质膜的横向组织和异质性在细胞功能 [ 211 ]。具体来说,分子几何形状可能会改变 LNP 脂质膜的刚度、曲率应变和可变形性 [ [212]、 [213]、 [214]、 [215]]。例如,帕特尔等人。探索了改变 LNP 中胆固醇衍生物的结构对细胞摄取和运输的影响,发现递送效率提高了 25 倍 [ 216 ],这可能是由 mRNA-LNP 内部和外部结构的形态变化引起的 [ 163 , 216 ]。此外,对 mRNA-LNP 内体逃逸的模拟表明,外周脂质向内体膜的横向扩散和尾部突出可能是实现内体融合的关键脂质动力学类型 [ 165 ]。最外层的脂质,PEG-脂质,可能对 LNP 膜的特性有其他修正。
PEG-脂质,通常是 LNP 配方中含量最低的脂质,以多种方式影响 LNP 的性质。首先,掺入的 PEG-脂质的量决定了粒径 [ 217 ],尽管过度的PEG 化会阻碍转染 [ 218 ]。其次,PEG-脂质可防止不稳定和聚集 [ 217 , 219 ]。第三,PEG-脂质可防止 LNP 的快速摄取,并减少血清蛋白的调理作用和网状内皮清除率,从而提高 LNP 循环寿命 [ 220 ]。最后,PEG 的亲水涂层可能有利于在粘性介质(如肺粘液)中导航 [ 116 , 221]。这些性质的程度取决于 PEG-脂质的化学结构,它包含亲水部分 (PEG) 和疏水部分 (脂质锚)。两者都可能影响 LNP 大小、脂质膜通透性和免疫反应 [ 66 , 222 , 223 ]。与 LNP 的其他成分不同,PEG-脂质旨在最终解离和脱落 PEG,从而控制细胞摄取的动力学。PEG 脱落对于防止潜在诱导 PEG 特异性抗体是必要的,这些抗体通过加速血液清除(ABC) 现象导致后续剂量的 PEG 化纳米材料的快速全身清除 [ 224]。ABC 的严重程度也可以通过调整 PEG 脂质结构来抑制,影响脱落的动力学和链的识别 [ 224 , 225 ]。这些发现表明,必须仔细调整 PEG-脂质的结构和数量以保持“隐形”效应。
mRNA-LNP 疫苗的长期储存是 LNP 生命周期中一个重要但尚未充分探索的部分。由于自组装结构通过弱分子间相互作用保持在一起,因此它们可能会发生结构变化以在长期储存中达到平衡。尚不清楚在储存中用于 mRNA 递送的自组装材料发生了哪些变化。然而,赵等人最近的一份报告。专门研究载有 mRNA 的脂质材料的长期储存条件表明,储存在水溶液中的类脂质纳米颗粒 (mRNA-LLN) 会发生尺寸变化并在体内失去其功效[ 226 ]。相比之下,冷冻干燥的 mRNA-LLN 在用 5%冷冻保护剂冻干时仍保持其功效20% 冷冻保护液冻干后,肝脏优先向脾脏摄取器官。这些发现支持自组装材料是对环境敏感的动态系统的假设,并强调需要进一步研究长期储存mRNA 疫苗的影响。
总之,为了实现高效、无毒和组织、器官或细胞选择性的 LNP 制剂,可以修改几乎无穷无尽的参数空间。尽管有一些结构标准(例如,可生物降解基团、可电离基团的 pKa 等)可用于设计用于 mRNA 递送的下一代脂质,但原子水平上的任何变化都可能需要额外的优化和大幅改进(或恶化)mRNA传递效率。mRNA 递送领域将极大地受益于深入的 SAR 和形态学研究,以建立通过 LNP 控制和靶向递送 mRNA 的指南。
6.2 . 基于聚合物的系统
复合物或基于聚合物的核酸载体是另一种流行的 mRNA 递送系统。通常,驱动 mRNA 复合物自组装的力与基于脂质的载体的情况相同——主要是静电吸引力(带正电荷的聚合物和带负电荷的核酸之间)、聚合物链之间的疏水相互作用,以及聚合物、溶剂和核酸货物之间的范德华力。然而,聚合物配方依赖于与基于脂质的配方不同的参数。聚合物可以在同一分子中包含不同的官能团,并且具有不同的长度和单体单元的排列[ 141]。因此,复合材料通常仅由一种可以同时获得多种功能的聚合物组成。尽管如此,聚合物材料,就像 LNP 一样,需要根据有效载荷优化聚合物成分 [ 227 ],这在很大程度上是经验性的。
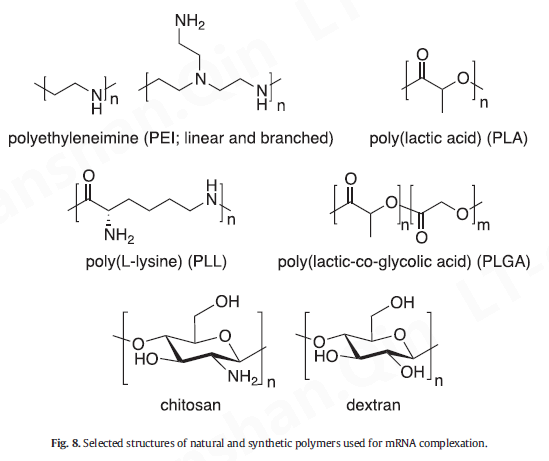
聚合基因传递系统最初与细胞毒性作斗争,部分是由于它们的阳离子电荷 [ 141 , 228 ]。基因传递的早期尝试主要基于三种类型的合成聚合物——聚(乙烯亚胺)(PEI)、聚(L-赖氨酸)(PLL)和聚(酰胺基胺)(PAMAM)(图8);然而,只有 PEI 被广泛用于 mRNA 疫苗递送 [ 150 ]。尽管优化的 PEI 结构提供了高基因转染效率,但其高阳离子电荷密度仍然会产生毒性问题 [ 111 ]。用 PEG 链和靶向部分装饰这些阳离子聚合物可减轻细胞毒性并大大改善其体外递送和体内[ 141、[228]、[229]、[230] ]。关于聚合物结构的另一个问题是生物降解性,这使得聚合物mRNA 载体的设计转向聚酯(例如,聚(β-氨基酯)(PBAE)、聚(胺-共-酯)(PACE)、聚(乳酸)酸)(PLA)、聚(乳酸-乙醇酸)(PLGA)和聚(2-二甲基氨基乙基甲基丙烯酸酯)(PDMAEMA))、天然聚合物(例如壳聚糖和葡聚糖)衍生物(图8)和其他可生物降解的结构 [ 4 , 229 , [231] , [232] , [233] ,[234] ]。例如,严等人。据报道,全身给药的基于聚酯的纳米颗粒优先将 mRNA 递送至肺部 [ 235 ],以及 Fornaguera 等人。证明寡肽末端修饰的 PBAE纳米载体在静脉注射后转染脾APC ,突出了它们作为 mRNA 疫苗的潜力[ 236 ]。
。。。。
参考资料
- Advanced Drug Delivery Reviews Volume 170, March 2021, Pages 83-112 。 Self-assembled mRNA vaccines。 https://www.sciencedirect.com/science/article/pii/S0169409X20302933?via%3Dihub
