【5.6.1.2】通过二级结构设计稳定信使RNA的理论基础--superfolder mRNA
设计的核心思想:
- 整体的AUP保持最低,不容易水解,mRNA更稳定,半衰期更长
- 5’AUP需要更高,特别是ATG起始的14个碱基, 这样翻译效率会更高。
我们定义了两个指标:
- 分子的总不配对概率(SUP,summed unpaired probability of a molecule)
- 平均不配对概率,它是按序列长度归一化的 SUP (AUP,the SUP normalized by sequence length)
具有低 AUP,我们称之为“superfolder”mRNA。
一、方法:
- LinearPartition-V ( 28 ) 用于计算 AUP
- RiboTree,蒙特卡洛树搜索算法,以随机最小化 mRNA 序列的 AUP。 ( https://eternagame.org/about/software )
- CDSFold ,通过运行具有不同最大碱基配对距离的 CDSfold 算法源代码 ( https://github.com/gterai/CDSfold , https://github.com/eternagame/CDSfold_SU )
- LinearDesign ( http://rna.baidu.com/ )
- Structure prediction and ensemble-based calculations were performed using LinearFold and LinearPartition with ViennaRNA and CONTRAfold parameters
- Secondary structure features were calculated from predicted MFE structures using RiboGraphViz (www.github.com/DasLab/RiboGraphViz).
- The codon adaptation index (CAI) was calculated as the geometric mean of the relative usage frequency of codons along the length of the coding region
- RNA secondary structures were visualized using draw_RNA (www.github.com/DasLab/draw_rna) and RiboGraphViz (www.github.com/DasLab/RiboGraphViz)
在线密码子优化个工具:
- Integrated DNA Technologies(IDT, https://www.idtdna.com/CodonOpt )
- Twist Biosciences( https://ecommerce.twistdna.com/app )
- GENEWIZ( https://clims4.genewiz.com/Toolbox/CodonOptimization )
二结论
2.1 RNA降解的生物物理模型
RNA 水解的速率是每个核苷酸处于未配对状态的平衡概率的属性,使其 3' 磷酸二酯键容易受到内联攻击构象的影响(图1B)。支持信息中介绍的该模型的完整推导假设配对或未配对核苷酸的降解速率遵循一级动力学(类似于为整合结构探测数据而开发的模型(31-33))。我们将 SUP 作为一种易于计算的可观察值引入,它与 RNA 分子的整体切割速率直接相关。

AUP 值是一个介于 0 和 1 之间的数字,它反映了 RNA 的整体“非结构化”,并说明了任何二级结构基序中的未配对区域。较低的值对应于未配对的较低概率,因此 RNA 分子不易降解。
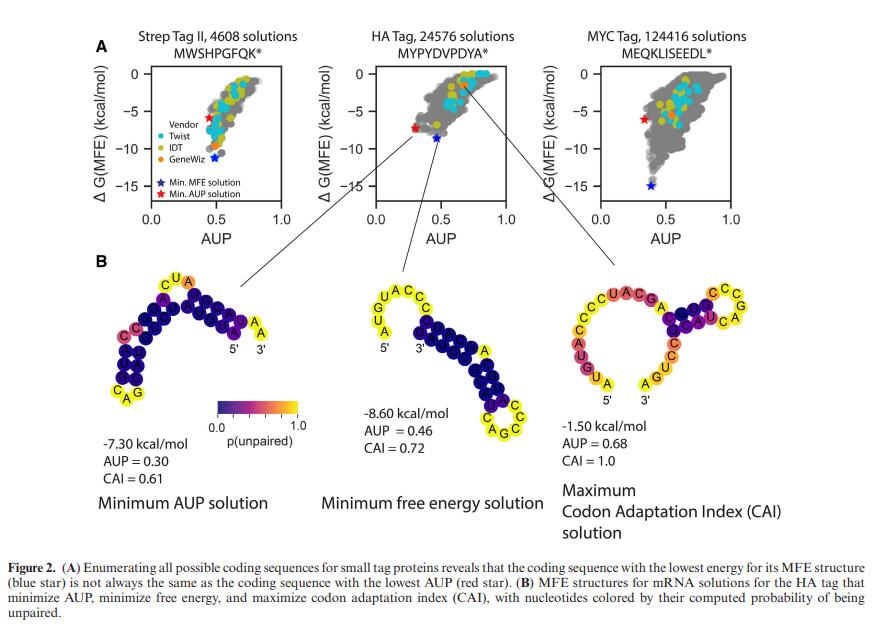
2.2 小型 mRNA 模型揭示了针对 SUP 优化的序列与针对密码子优化或最小折叠自由能优化的序列之间的差异
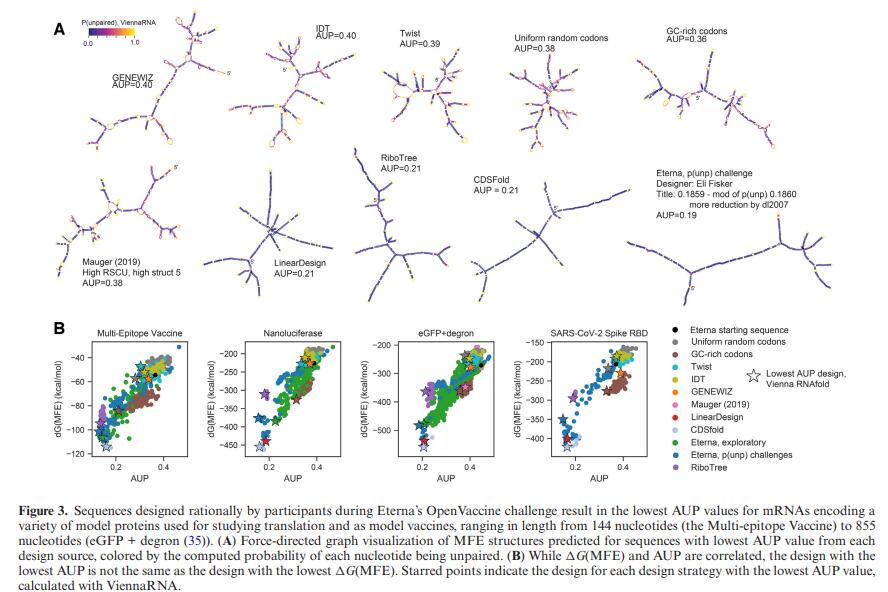
2.3 对于长 mRNA 构建体,AUP 可降低两倍
2.4 与翻译和免疫原性功能相关的特性的多样性
在确定设计具有减少的 AUP 的 mRNA 序列的可行性后,我们希望确定这些序列是否可用于翻译以及预防或引发先天免疫反应。
我们首先描述了跨设计方法的序列的 CAI ( 30 ),因为该措施与提高翻译效率有关 ( 40-44 )。我们发现,在所有 mRNA 设计挑战中,最小 AUP 序列的 CAI 值始终大于 0.7(图4A)。另一个被假设影响蛋白质翻译效率的设计特征是 CDS 直接暴露在起始密码子的上游 ( 45 , 46 )。我们计算了前 14 个核苷酸 ( 45 ) 的平均未配对概率,称为 AUP init,14,在我们的模型 UTR 来自人类血红蛋白亚基 β (HBB) 的情况下。更高的 AUP 值init,14表明更多暴露的核糖体起始位点,预计与更高的翻译效率相关。我们发现了一系列 AUP init,低 AUP 序列可能有 14 个值(图4B)。这些分析表明,设计可翻译的低 AUP 序列是可行的,正如 CAI 和 AUP init 的可用指标所评估的那样,14。
另一个重要的考虑因素是 mRNA 治疗剂从识别双链 RNA 螺旋的途径中引发免疫原性反应的可能性 ( 47-49 )。我们发现特征序列中没有一个包含长度超过 33 个碱基对的 Watson-Crick 螺旋,该长度已被发现是在蛋白激酶 R (PKR) 感应后导致细胞 mRNA 翻译全局关闭的最小长度 ( 50 ),不超过 112 bp,观察到的长度可刺激视黄酸诱导基因 I (RIG-I) 样受体黑色素瘤分化相关蛋白 5 (MDA5) ( 48 ) 的协同结合(图4C)。然而,PKR 已被表征为结合含有凸起的 dsRNA,并在 A 型 dsRNA 的 ~2 倍亲和力范围内错配一个或两个核苷酸长 ( 51 )。因此,我们还计算了考虑单核苷酸和双核苷酸凸起以及 1 × 1 和 2 × 2 核苷酸内部环的所有设计构建体的最大 dsRNA 长度(补充图S6)。考虑到这些缺陷时,许多低 AUP 结构的最大 dsRNA 长度大于 33 bp,但仍短于 112 bp。如果用未修饰的核苷酸合成,dsRNA 结合蛋白可能以降低的亲和力结合这些构建体。一般而言,低 AUP 构建体可实现的最大 dsRNA 长度的多样性表明,可能会实现不太剧烈的先天免疫反应,并且该反应可能是可调节的,这取决于这种反应是否可取(mRNA 疫苗)(例如抗免疫 mRNA 疗法)。
参考资料
- Theoretical basis for stabilizing messenger RNA through secondary structure design。 https://academic.oup.com/nar/article/49/18/10604/6370252
